Note: Descriptions are shown in the official language in which they were submitted.
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99I00656
SARA PROTEINS w
Field of the Invention
The invention relates to a family of proteins, the SARA proteins, which
bind to receptor-regulated Smad proteins and are involved in appropriate
localization of these Smad proteins for receptor activation.
Background of the Invention
The Transforming Growth Factor-beta (TGF(3) superfamily, whose
members include TGF~is, activins and bone morphogenetic proteins (BMPs),
have wide ranging effects on cells of diverse origins (Attisano and Wrana,
1998;
Heldin et al., 1997; Kretzschmar and Massague, 1998). Signaling by these
secreted factors is initiated upon interaction with a family of cell-surface
transmembrane serine/threonine kinases, known as type I and type II receptors.
Ligand induces formation of a typel/typell heteromeric complex which permits
the constitutively active type II receptor to phosphorylate, and thereby
activate,
the type I receptor (Wrana et al., 1994). This activated type I receptor then
propagates the signal to a family of intracellular signaling mediators known
as
Smads (Attisano and Wrana, 1998; Heldin et al., 1997; Kretzschmar and
Massague, 1998).
The first members of the Smad family identified in invertebrates were the
Drosophila MAD and the C. elegans sma genes (sma-2, sma-3 and sma-4; Savage
et al., 1996; Sekelsky et al., 1995). Currently, the family includes
additional
invertebrate Smads, as well as nine vertebrate members, Smad1 through 9
25 (Attisano and Wrana, 1998; Heldin et al., 1997; Kretzschmar and Massague,
1998). Smad proteins contain two conserved amino (MH1) and carboxy (MH2)
terminal regions separated by a more divergent linker region. !n general, Smad
proteins can be subdivided into three groups; the receptor-regulated Smads,
which include Smad 1, 2, 3, 5 and 8, Mad, sma-2 and sma-3; the common
Smads, Smad4 and Medea, and the antagonistic Smads, which include Smad6, 7
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
2
and 9, DAD and daf-3 (Heldin et al., 1997; Nakayama et al., 1998; Patterson et
al., 1997).
Numerous studies with vertebrate Smad proteins have provided insights
into the differential functions of these proteins in mediating signaling.
Receptor-
regulated Smads are direct substrates of specific type I receptors and the
proteins
are phosphorylated on the last two serines at the carboxy-terminus within a
highly conserved SSXS motif (Abdollah et al., 1997; Kretzschmar et al., 1997;
Liu et al., 19976; Macias-Silva et al., 1996; Souchelnytskyi et al., 1997).
Interestingly, Smad2 and Smad3 are substrates of TGF/3 or activin receptors
and
10 mediate signaling by these ligands (Liu et al., 19976; Macias-Sifva et al.,
1996;
Nakao et al., 1997a), whereas Smadl, S and 8 appear to be targets of BMP
receptors and thereby propagate BMP signals (Chen et al., 19976; Hoodless et
al., 1995; Kretzschmar et al., 1997; Nishimura et al., 1998). Once
phosphorylated, these Smads bind to the common Smad, Smad4, which lacks
the carboxy-terminal phosphorylation site and is not a target for receptor
phosphorylation (Lagna et al., 1996; Zhang et al., 1997). Heteromeric
complexes of the receptor-regulated Smad and Smad4 translocate to the nucleus
where they function to regulate the transcriptiona) activation of specific
target
genes. The antagonist Smads, Smad6, 7 and 9 appear to function by blocking
20 ligand-dependent signaling by preventing access of receptor-regulated Smads
to
the type I receptor or possibly by blocking formation of heteromeric complexes
with Smad4 (reviewed in Heldin et al., 1997).
Analysis of the nuclear function of Smads has demonstrated that Smads
can act as transcriptional activators and that some Smads, including
Drosophila
Mad, and the vertebrate Smad3 and Smad4, can bind directly to DNA, albeit at
relatively low specificity and affinity (Dennler et al., 1998; Kim et al.,
1997;
Labbe et al., 1998; Yingling et al., 1997; Zawel et al., 1998).
Localization of Smads is critical in controlling their activity and Smad
phosphorylation by the type I receptor regulates Smad activity by inducing
nuclear accumulation (Attisano and Wrana, 1998; Heldin et al., 1997;
Kretzschmar and Massague, 1998). However, little is known about how Smad
CA 02335346 2001-O1-18
WO 00/05360 PC'T/CA99/00656
3
localization is controlled prior to phosphorylation and how this might
function
in modulating receptor interactions with its Smad substrates.
Summary of the Invention
S Smad proteins (Smads) transmit signals from transmembrane ser/thr kinase
receptors to the nucleus. Mammalian and non-mammalian proteins have been
identified which interact directly with Smads and are designated the Smad
Anchor for Receptor Activation or SARA proteins.
The invention provides cDNA sequences encoding this previously
10 undescribed family of SARA proteins which bind to receptor-regulated Smad
proteins and ensure appropriate localization of these Smad proteins for
activation by a Type I receptor of a TGF~3, activin or BMP signaling pathway.
For example, TGF~3 signaling induces dissociation of Smad2 or Smad3
from a SARA protein with concomitant formation of Smad2/Smad4 or
15 Smad3/Smad4 complexes and nuclear translocation. In the absence of
signaling,
SARA functions to recruit a particular Smad (eg. Smad2 or Smad3) to distinct
subcellufar sites in the cell and interacts with the TGF~i superfamily
receptor
complex in cooperation with the particular receptor regulated Smad. Mutations
in hSARAI that cause mislocalization of Smad2, and interfere with receptor
20 association, inhibit receptor-dependent transcriptional responses,
indicating that
regulation of Smad localization is essential for TGF(i superfamily signaling.
The
invention provides a novel component of the signal transduction pathway that
functions to anchor Smads to specific subcellular sites for activation by the
Type
I receptor of the TGF~i, activin or BMP signaling pathways.
25 The SARA proteins are characterised by the presence of three domains, a
double zinc finger or FYVE domain responsible for the subcellular localisation
of
the SARA protein or SARA-Smad complex, a Smad-binding domain which
mediates the interaction or binding of one or more species of Smad protein and
a carboxy terminal domain which mediates association with the TGF~3
30 superfamily receptor. The FYVE domain may bind phosphatidyl inositol-3-
phosphate.
CA 02335346 2001-O1-18
WO 00/05360 PC'T/CA99/00656
4
In accordance with one embodiment, the invention provides isolated
polynucleotides comprising nucleotide sequences encoding SARA proteins.
fn accordance with a further series of embodiments, the invention
provides an isolated pofynucleotide selected from the group consisting of
(a) a nucleotide sequence encoding a human SARA protein;
(b) a nucleotide sequence encoding a mammalian SARA protein;
(c) a nucleotide sequence encoding a non-mammalian SARA
protein;
(d) a nucleotide sequence encoding the human SARA amino acid
sequence of Table 2 (hSARAI: Sequence ID N0:2);
(e) a nucleotide sequence encoding the human SARA amino acid
sequence of Table 4 (hSARA2: Sequence ID N0:4);
(f) a nucleotide sequence encoding the Xenopus SARA amino acid
sequence of Table 6 (XSARA1: Sequence ID N0:6);
(g) a nucleotide sequence encoding the Xenopus SARA amino acid
sequence of Table 8 (XSARA2: Sequence ID N0:8).
In accordance with a further embodiment, the invention provides the
nucleotide sequences of Table 1 (human SARA1 or hSARAI), Table 3 (human
SARA2 or hSARA2), Table 5 (Xenopus SARA1 or XSARA1 ) and Table 7 (Xenopus
SARA2 or XSARA2).
In accordance with a further embodiment, the invention provides
recombinant vectors including the polynucleotides disclosed herein and host
cells transformed with these vectors.
The invention further provides a method for producing SARA proteins,
comprising culturing such host cells to permit expression of a SARA protein-
encoding polynucleotide and production of the protein.
The invention also includes polynucleotides which are complementary to
the disclosed nucleotide sequences, polynucleotides which hybridize to these
sequences under high stringency and degeneracy equivalents of these
sequences.
CA 02335346 2001-O1-18
WO 00/05360 PCTlCA99/00656
In accordance with a further embodiment, the invention provides
antisense molecules which may be used to prevent expression of a SARA
protein. Such antisense molecules can be synthesised by methods known to
those skilled in the art and include phosphorothioates and similar compounds.
5 The invention further includes polymorphisms and alternatively spliced
versions of the disclosed SARA genes and proteins wherein nucleotide or amino
acid substitutions or deletions do not substantially affect the functioning of
the
gene or its encoded protein.
The invention also enables the identification and isolation of allelic
variants or homologues of the described SARA genes, and their corresponding
proteins, using standard hybridisation screening or PCR techniques.
The invention provides a method for identifying allelic variants or
homologues of the described SARA genes, comprising
choosing a nucleic acid probe or primer capable of hybridizing to a SARA
7 5 gene sequence under stringent hybridisation conditions;
mixing the probe or primer with a sample of nucleic acids which may
contain a nucleic acid corresponding to the variant or homologue; and
detecting hybridisation of the probe or primer to the nucleic acid
corresponding to the variant or homologue.
In accordance with a further embodiment, the invention provides
fragments of the disclosed polynucieotides, such as polynucleotides of at
least
10, preferably 15, more preferably 20 consecutive nucleotides of the disclosed
polynucleotide sequences. These fragments are useful as probes and PCR
primers or for encoding fragments, functional domains or antigenic
determinants
of SARA proteins.
In accordance with a further embodiment, the invention provides
substantially purified SARA proteins, including the proteins of Table 2
(hSARAI),
Table 4 (hSARA2), Table 6 (XSARA1) and Table 8 (XSARA2).
in accordance with one embodiment, a SARA protein has a FYVE domain,
a Smad binding domain (SBD) and an amino acid sequence having at least
50°/°
overall identity with the amino acid sequence of hSARAI (Sequence !D N0:2).
CA 02335346 2001-O1-18
1~V0 00/05360 PCT/CA99/00656
6
In accordance with a preferred embodiment, a SARA protein has a FYVE
domain having at least 65% identity of amino acid sequence with the FYVE
domain of hSARAI and a C-terminal sequence of 550 consecutive amino acids
which have at least 50% identity with the C-terminal 550 amino acid residues
of
hSARAI.
in accordance with a more preferred embodiment, a SARA protein has a
FYVE domain having at least 65°/° identity of amino acid
sequence with the
FYVE domain of hSARAI and wherein the portion of the SBD corresponding to
amino acid residues 721 to 740 of hSARAI has at least 80°l0 identity
with that
portion of hSARAI.
The invention further provides a method for producing antibodies which
selectively bind to a SARA protein comprising the steps of
administering an immunogenically effective amount of a SARA
immunogen to an animal;
allowing the animal to produce antibodies to the immunogen; and
obtaining the antibodies from the animal or from a cell culture derived
therefrom.
The invention further provides substantially pure antibodies which bind
selectively to an antigenic determinant of a SARA protein. The antibodies of
the
invention include pofyclonal antibodies, monoclonal antibodies and single
chain
antibodies.
The invention includes analogues of the disclosed protein sequences,
having conservative amino acid substitutions therein. The invention also
includes fragments of the disclosed protein sequences, such as peptides of at
least 6, preferably 10, more preferably 20 consecutive amino acids of the
disclosed protein sequences.
The invention further provides polypeptides comprising at least one
functional domain or at least an antigenic determinant of a SARA protein.
In accordance with a further embodiment, the invention provides
peptides which comprise SARA protein Smad binding domains and
polynucleotides which encode such peptides.
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
7
In accordance with a further embodiment, the invention provides a Smad
binding domain peptide selected from the group consisting of
(a) SASSQSPNPNNPAEYCST1PPLQQAQASGALSSPPPTVMVPVGV
LKHPGAEVAQPREQRRVWFADGILPNGEVADAAKLTMNGTSS; and
(b) amino acids 589 to 672 of the XSARA1 sequence of Table 9.
The invention includes fragments and variants of these Smad binding
domain peptides which retain the ability to bind a Smad protein.
In accordance with a further embodiment, the invention provides
peptides which comprise SARA protein FYVE domains and polynucleotides
which encode such peptides.
In accordance with a further embodiment, the invention provides a FYVE
domain peptide selected from the group consisting of
(a) amino acids 587 to 655 of the hSARAI sequence of Table 9;
(b) amino acids 510 to 578 of the XSARA1 sequence of Table 9; and
(c) the consensus amino acid sequence of Table 10.
The invention includes fragments and variants of these FYVE domain
peptides which retain the function of the parent peptide.
In accordance with a further embodiment, the invention provides
peptides which comprise SARA protein TGF(3 receptor interacting domains and
polynucleotides which encode such peptides.
fn accordance with a further embodiment, the invention provides a TGF~i
receptor interacting domain peptide comprising amino acids 751 to 1323 of the
hSARAI sequence of Table 9.
The invention includes fragments and variants of these TGF~3 receptor
binding domain peptides which retain the binding ability of the parent
peptide.
The invention further provides methods for modulating signaling by
members of the TGF(3 superfamily which signal through pathways which involve
a SARA protein.
Modulation of signaling by a TGF~i superfamily member through such a
pathway may be effected, for example, by increasing or reducing the binding of
the SARA protein involved in the pathway with its binding partner.
CA 02335346 2001-O1-18
WO 00/05360 PC'T/CA99/00656
8
In accordance with a further embodiment, TGF(3 superfamily signaling,
including TGF(3 signaling, by a pathway involving a SARA protein described
herein may be modulated by modulating the binding of the SARA protein to a
Smad binding partner, by modulating the binding of its FYVE domain to its
binding partner or by modulating the binding of the SARA protein to a TGF~3
superfamily receptor, such as the TGF(3 receptor.
For example, the binding of a SARA protein to a Smad binding partner
may be inhibited by a deletion mutant of the protein lacking either the SBD
domain or the FYVE domain or by the SARA protein Smad binding domain
10 peptides or FYVE domain peptides described herein, and effective fragments
or
variants thereof. The binding of a SARA protein to a TGF~3 superfamily
receptor
may be inhibited by a deletion mutant of the protein lacking a C terminal
portion
or by the SARA protein TGF~i receptor binding domain peptides described
herein, and effective fragments and variants thereof.
15 In accordance with a further embodiment, TGF~3 superfamily signaling,
including TGF(3 signaling, by a pathway involving a SARA protein may be
modulated by modulating the binding of the SARA protein FYVE domain to
phosphatidyl inositol-3-phosphate, by increasing or decreasing the
availability of
phosphatidyl inositol-3-phosphate or by administration of agonists or
antagonists
20 of phosphatidyl inositol-3-phosphate kinase.
The invention also provides a method of modulating a TGF~i superfamily
signaling pathway involving phosphatidyl inositol-3-phosphate, including a
TGF(3 signaling pathway, by increasing or decreasing the availability of SARA
protein or by modulating the function of SARA protein.
25 The invention further provides methods for preventing or treating diseases
characterised by an abnormality in a TGF(3 superfamily member signaling
pathway which involves a SARA protein, by modulating signaling in the
pathway, as described above.
TGF~3 signaling is important in wound healing, and excessive signaling is
30 associated with scarring, with arthritis and with fibrosis in numerous
diseases,
including fibrosis of the liver and kidney. TGF~i signaling is also involved
in
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
9
modulating inflammatory and immune responses and can contribute to tumour
progression.
The invention thus provides methods for modulating TGF(3-dependent
cell proliferation or fibrogenesis.
The BMP signaling pathways are important in tissue morphogenesis and
in protecting tissues and restoring or regenerating tissues after tissue
damage, for
example in bone, kidney, liver and neuronal tissue (see, for example, (Reddy,
A.H. (1998), Nature Biotechnology, v. 16, pp. 247-252).
The invention further provides methods for modulating BMP-dependent
phenotypic marker expression by modulating the interactions of SARA proteins
involved in these BMP signaling pathways.
In accordance with a further embodiment, modified versions of a SARA
protein may be provided as dominant-negatives that block TGF(3 superfamily
signaling. These modified versions of SARA could, for example, lack the Smad
binding domain and thereby prevent recruitment of Smad or could lack the FYVE
domain and thereby inhibit signaling by interfering with translocation.
These modified versions of SARA may be provided by gene therapy, for
example using transducing viral vectors. Expression may be driven by inclusion
in the vector of a promoter specific for a selected target cell type. Many
examples of such specific promoters are known to those skilled in the art.
In a further embodiment, a normal version of a SARA protein such as
hSARAI could be provided by gene therapy to restore function in a disease
wherein SARA is mutated or non-functional.
In a further embodiment, the invention provides a pharmaceutical
composition comprising a purified SARA protein as active ingredient.
In accordance with a further embodiment, the invention provides non-
human transgenic animals and methods for the production of non-human
transgenic animals which afford models for further study of the SARA system
and
tools for screening of candidate compounds as therapeutics. For example, knock
out animals, such as mice, may be produced with deletion of a SARA gene.
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
These animals may be examined for phenotypic changes and used to screen
candidate compounds for effectiveness to reverse these changes.
In a further example, transgenic animals may be produced expressing a
dominant negative mutant of a SARA protein, as described above, either
5 generally or in specific targeted tissues.
The invention provides many targets for the development of small
molecule drugs, including peptides and peptidomimetic drugs, to interfere with
the interaction of the various binding partners described herein and thereby
modulate signaling by members of the TGF~i superfamily, including TGF~ and
10 BMPs
The invention further provides methods for screening candidate
compounds to identify those able to modulate signaling by a member of the
TGF~i superfamily through a pathway involving a SARA protein.
For example, the invention provides screening methods for compounds
able to bind to a SARA protein which are therefore candidates for modifying
the
activity of the SARA protein. Various suitable screening methods are known to
those in the art, including immobilization of a SARA protein on a substrate
and
exposure of the bound SARA protein to candidate compounds, followed by
elution of compounds which have bound to the SARA protein. The methods
used to characterise the binding interactions of the SARA proteins disclosed
herein, as fully described in the examples herein, may also be used to screen
for
compounds which are agonists or antagonists of the binding of a SARA protein.
This invention also provides methods of screening for compounds which
modulate TGF(3 superfamily signaling by detecting an alteration in the
phosphorylation state of a SARA protein.
In accordance with a further embodiment, the invention provides a
method for reducing or preventing TGF~3, activin or BMP signaling by
inhibiting
the activity of SARA. SARA activity may be inhibited by use of an antisense
sequence to the SARA gene or by mutation of the SARA gene.
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
11
Summary of the Drawings
Certain embodiments of the invention are described, reference being
made to the accompanying drawings, wherein:
Figure 1 (top panel) shows interaction of full length hSARAI with
bacterially expressed Smads. Full length SARA protein was produced in an in
vitro transcription/translation system in the presence of [35S]methionine and
was
incubated with glutathione-sepharose beads coated with bacterially-expressed
GST fusion proteins of the indicated Smads or Smad2 subdomains. Bound
material was resolved by SDS-PAGE and visualized by autoradiography.
Migration of full length hSARAI, and a translation product that initiates from
an
internal methionine located upstream of the Smad binding domain (asterisk) are
indicated. The presence of approximately equivalent amounts of GST fusion
proteins was confirmed by SDS-PAGE and coomassie staining of a protein
aliquot (bottom panel).
Figure 2 shows interaction of hSARA with Smads in mammalian cells.
COS cells were transfected with Flag-tagged hSARAI (Flag-SARA) either alone or
together with the indicated Myc-tagged Smad constructs. For Smad6, an
alternative version lacking the MH 1 domain was used (Topper et al., 1997).
Cell
lysates were subjected to an anti-Flag immunoprecipitation and coprecipitating
Smads detected by immunoblotting with anti-Myc antibodies. The migration of
anti-Flag heavy and light chains (IgG) are marked. To confirm efficient
expression of hSARAI and the Smads, aliquots of total cell lysates were
immunoblotted with the anti-Flag and anti-Myc antibodies (bottom panel). The
migrations of hSARAI and the Smads are indicated.
Figures 3-6 show immunoblots of lysates from COS cells transiently
transfected with various combinations of Flag or Myc-tagged hSARAI, wild type
(WT) or mutant (2SA) Myc or Flag-tagged Smad2, Smad4/HA and wild type (WT)
or constitutively active (A) T(3R1/HA, cell lysates being subjected to
immunoprecipitation with anti-Flag or anti-Myc antibodies, as indicated.
Confirmation of protein expression was performed by immunoblotting total cell
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
12
lysates prepared in parallel for the indicated tagged protein (totals, bottom
panels).
Figure 3: Transfected cells were metabolically labelled with [3zP]PO4 and
cell lysates subjected to immunoprecipitation with anti-Flag antibodies for
visualization of hSARAI phosphorylation (top panel) or with anti-Myc
antibodies
for Smad2 phosphorylation (middle panel). Immunoprecipitates were resolved
by SDS-PAGE and visualized by autoradiography. The migrations of hSARAI
and Smad2 are indicated.
Figure 4: Lysates from transiently transfected COS cells were subjected to
immunoprecipitation with anti-Flag antibodies and Smad2 bound to hSARAI
was analyzed by immunoblotting with anti-Myc antibodies (IP: a-flag; blot: a-
Myc).
Figure 5: Lysates from transiently transfected COS cells were subjected to
immunoprecipitation with anti-Flag antibodies and Smad2 bound to hSARAI
was analyzed by immunoblotting with anti-Myc antibodies (IP: a-flag, blot: a-
Myc). Partial dissociation of hSARAI/Smad2 complexes induced by TGF(3
signaling was enhanced by expression of Smad4.
Figure 6: Cell lysates from transiently transfected COS cells were
subjected to immunoprecipitation with anti-Fiag antibodies directed towards
Smad2. Immunoprecipitates were then immunobfotted using anti-Myc or anti-
HA antibodies which recognize hSARAI or Smad4, respectively.
Coprecipitating SARA (a-myc blot) and Smad4 (a-HA blot) are indicated.
Figure 7, panels A to E, shows photomicrographs of Mv1 Lu cells
transiently transfected with various combinations of Flag-Smad2, Myc-hSARAI,
and constitutively active T~iRI (T(3R1*) as indicated (Tx). hSARA was
visualized
with the polycfonal Myc A14 antibody and Texas-Red conjugated goat-anti-
rabbit IgG (red) and Smad2 was detected with an anti-Ffag M2 monoclonal
antibody followed by FITC-conjugated goat anti-mouse IgG (green). The
subcellular localization of the expressed proteins was visualized by
immunofiuorescence and confocal microscopy.
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
13
Panels A, B, C, Mv1 Lu cells singly transfected with hSARA1 (A) or Smad2
(B) are shown. Cotransfection of Smad2 with the constitutively active T~iRI
(T(3R1*) results in its accumulation in the nucleus (C).
Panel D, Mv1 Lu cells were transfected with hSARAI and Smad2 and the
localization of hSARAI (red, left photo) and Smad2 (green, centre photo) is
shown. Colocalization of SARA and Smad2 is shown (right photo) and appears
as yellow.
Pane) E, Mv1 Lu cells were transfected with hSARAI, Smad2 and activated
T~iRI (T(3R1*) and the localization of hSARA (red, left photo) and Smad2
(green,
centre photo) is shown. Colocalization of SARA and Smad2 is indicated (right
photo). Note the shift to an orangy-red colour in the punctate spots and an
intensification of Smad2 nuclear staining, indicative of dissociation of Smad2
from SARA and nuclear translocation.
Figure 7, panel F, shows photomicrographs of Mv1 Lu cells stained with
rabbit, polyclonal anti-SARA antibody (left photo, green), goat, polyclonal
anti-
Smad 2/3 antibody (centre photo, red) and with both antibodies (right photo,
yellow), showing co-localization of hSARAI and Smad2.
Figure 8A shows photomicrographs of Mv1 Lu cells transfected with either
hSARAI alone (panel i), T[3R11 alone (panel ii) or hSARAI and T(3R11 together
(panel iii), then treated with TGF(3 and the localization of hSARAI (red) and
T~iRll (green) determined by immunofluorescence and confocal microscopy. In
cells coexpressing hSARAI and T~iRll, superimposing the staining revealed
colocalization of the proteins as indicated by yellow staining in panel iii.
Figure 8B shows affinity labelling of COS cells transiently transfected with
various combinations of Flag-hSARAI, Myc-Smad2, wild type (WT) T~iRll and
either wild type or kinase-deficient (KR) versions of T~3R1. Cells were
affinity-
labelled with ['zSIJTGF~3 and lysates immunoprecipitated with anti-Flag
antibodies. Coprecipitating receptor complexes were visualized by SDS-PAGE
and autoradiography. Equivalent receptor expression was confirmed by
visualizing aliquots of total cell lysates (bottom panel).
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
14
Figure 9A shows COS cells transiently transfected with wild type T(3R11
and kinase-deficient T(3R1 and various combinations of wild type Flag-hSARAI
(WT), a mutant version lacking the Smad2 binding domain (~SBD) and Myc-
Smad2. The amount of receptor bound to SARA was determined by anti-Flag
immunoprecipitation followed by gamma counting. Data is plotted as the
average of three experiments ~ S.D. Protein expression was analyzed by
immunoblotting aliquots of total cell lysates and the results from a
representative
experiment are shown (bottom panel).
Figure 9B shows COS cells transiently transfected with wild type T~3R11
and kinase-deficient T~iRI and Flag-tagged wild type (WT) or mutant versions
of
hSARAI with (black bars) or without (open bars) Myc-Smad2. The amount of
receptor bound to hSARAI was determined by anti-Flag immunoprecipitation
followed by gamma counting. Protein expression was analyzed by
immunoblotting aliquots of total cell lysates (bottom panel).
Figure 10 is a schematic representation of mutant versions of SARA. The
FYVE domain (shaded bar) and the Smad binding domain, SBD (striped bar), are
indicated. COS cells transiently transfected with Fiag-hSARAI and Myc-Smad2
were immunoprecipitated with anti-Flag antibodies followed by immunoblotting
with anti-Myc antibodies. The presence (+) or absence (-) of a hSARAI/Smad2
interaction is indicated (Smad2 interaction). Mutants used for the subsequent
localization study are marked on the left (i-vi).
Figure 11 A shows an immunoblot of lysates from COS cells expressing
Flag-tagged Smad2 or Smad3 incubated with GST alone or with GST-hSARAI
(665-750), which corresponds to the SBD; bound proteins were immunoblotted
using anti-Flag antibodies. The presence of Smad2 and Smad3 bound to GST-
hSARAI (665-750) is indicated.
Figure 11 B shows an immunoblot of lysates, from COS cells expressing
Flag-tagged Smad2 together with wild type (WT) or activated (A) type I
receptor,
incubated with GST-hSARAI (665-750) (GST-SBD) and immunoblotted with anti-
Flag antibodies. The expression levels of Smad2, each receptor and GST-
CA 02335346 2001-O1-18
WO 00/05360 PC1'/CA99/00656
hSARAI (665-750) were determined by immunoblotting aliquots of total cell
lysates.
Figure 12 shows the subcellular localization of hSARAI mutants. Mv1 Lu
cells were transiently transfected with wild type (panel i) or mutant versions
of
5 Flag-hSARAI (panels ii-viii, as marked on the left in Figure 10). Proteins
were
visualized by immunofluorescence and confocal microscopy using a monoclonal
anti-Fiag M2 monoclonal antibody followed by FITC-conjugated goat anti-mouse
IgG.
Figure 13 shows photomicrographs of Mv1 Lu cells transiently transfected
10 with mutant versions of Myc-hSARAI and Flag-Smad2 (panel A) or with wild
type Myc-hSARAI, HA-Smad2 and mutant versions of hSARAI (panel B).
Protein subcelfular localization was visualized by immunofluorescence and
confocal microscopy. hSARAI was visualized with the polyclonal Myc A14
antibody and FITC-conjugated goat anti-rabbit IgG (green), while Smad2 was
15 detected with monoclonal antibodies followed by Texas Red-conjugated goat
anti-mouse IgG (red). In B, overlaying the images reveals mislocalization of
Smad2 as green speckles of SARA over red, diffuse Smad2 staining (panels ii
and
iii) and colocalization of hSARAI and Smad2 appears as yellow spots (panels i
and iv).
Figure 14 shows luciferase activity of Mv1 Lu cells transfected with 3TP-
lux alone or together with the indicated amounts of wild type (WT) or mutant
(01-664 or D1-704) versions of hSARAI and incubated in the presence (black
bars) or absence (open bars) of TGF~i. Luciferase activity was normalized to
~3-
galactosidase activity and is plotted as the mean ~S.D. of triplicates from a
representative experiment.
Figure 15 shows luciferase activity of HepG2 cells transfected with ARE-
Lux alone (v), or ARE-Lux and FAST2 alone or together with the indicated
amounts of wild type (WT) or mutant versions of hSARAI. Transfected cells
were incubated in the presence (black bars) or absence (open bars) of TGF(3
and
luciferase activity was normalized to (3-galactosidase activity and is plotted
as the
mean ~S.D. of triplicates from a representative experiment.
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
16
Figure 16 shows a Northern blot of expression of hSARAI (upper panel)
and Smad2 (lower panel) in the indicated tissues.
Figure i 7 shows an immunoblot of a HepG2 iysate immunoprecipitated
(IP) with preimmune serum (PI), anti-hSARAI polyclonal antibody (SARA) with
and without pretreatment with TGF~i (- and +), or N19 anti-Smad2/3 antibody
(S2), followed by immunoblotting with an anti-Smad2 antibody. The migration
position of Smad2 is indicated (Smad2).
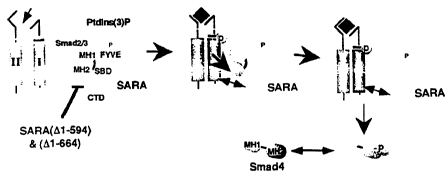
Figure 18 shows a diagram of a model of the interaction of a SARA
protein with a receptor regulated Smad, as exemplified by the interaction of
hSARAI.
Detailed Description of the Invention
This invention provides a family of proteins that play key roles in TGF-(3,
activin and bone morphogenetic protein (BMP) signal transduction pathways. In
particular, the proteins of this family interact with specific Smad proteins
to
modulate signal transduction. These proteins are therefore designated as "Smad
Anchor for Receptor Activation" or "SARA" proteins. SARA proteins are
characterised by three distinct domains (1) a double zinc finger or FYVE
domain
responsible for the subcellular localization of the SARA protein or SARA-Smad
complex, possibly through its association with Ptdlns(3)P, (2) a Smad binding
domain ("SBD") which mediates the interaction or binding of one or more
species of Smad protein with the particular member of the SARA family and (3)
a
carboxy terminal domain which mediates interaction of SARA with members of
the TGF(3 superfamily of receptors.
FYVE domains have been identified in a number of unrelated signaling
molecules that include FGD1, a putative guanine exchange factor for Rho/Rac
that is mutated in faciogential dysplasia, the HGF receptor substrate Hrs-1
and its
homolog Hrs-2, EEA1, a protein involved in formation of the early endosome
and the yeast proteins FAB1, VPS27 and VAC1 (reviewed in Wiedemann and
Cockcroft, 1998). Recently, analysis of a number of FYVE domains from yeast
and mammals has revealed that this motif binds phosphatidyl inositol-3
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
17
phosphate (Ptdins(3)P) with high specificity and thus represents a novel
signaling
module that can mediate protein interaction with membranes (Burd and Emr,
1998; Gaullier et al., 1998; Patki et al., 1998; Simonsen et al., 1998;
Wiedemann and Cockcroft, 1998). Comparison of the FYVE domains from the
vertebrate proteins with that from SARA revealed extensive conservation of
residues throughout the domain (Table 10). Thus, SARA contains a FYVE
domain that may function to bind Ptdlns(3)P, which has been implicated in
intracellular vesicle transport.
For example, deletion of the FYVE domain in hSARAI causes
mislocalization of Smad2 or Smad3, interferes with TGF(3 receptor interaction
and inhibits TGF(3-dependent transcriptional responses.
Thus, the SARA proteins of the invention define a component of TGF~
superfamily signaling that fulfills an essential role in anchoring receptor
regulated Smads to specific subcellular domains for activation by a TGF/3
superfamily receptor.
Cloned DNA coding sequences and corresponding amino acid sequences
for representative human and Xenopus SARA protein family members are shown
in the Tables, as follows:
Tables 1 and 2 - human SARA1 (hSARAI) cDNA (Sequence ID N0:1)
and amino acid sequence (Sequence ID N0:2) respectively;
Tables 3 and 4 - human SARA2 (hSARA2) cDNA (Sequence ID N0:3)
and amino acid sequence (Sequence ID N0:4) respectively;
Tables 5 and 6 - Xenopus SARA1 (XSARAi ) cDNA (Sequence ID NO:S)
and amino acid sequence (Sequence ID N0:6) respectively; and
Tables 7 and 8 - Xenopus SARA2 (XSARA2) cDNA (Sequence ID N0:7)
and amino acid sequence (Sequence ID N0:8) respectively.
Table 9 shows a comparison of the amino acid sequences of XSARA1 and
hSARAI . Identical residues (dark grey) and conservative changes (fight grey),
the FYVE domain (solid underline) and the Smad binding domain (dashed
underline) are indicated. The sequences in XSARA1 used to design degenerate
PCR primers for identifying hSARAI are shown (arrows). The amino-terminal
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99100656
18
end of the partial Xenopus cDNA obtained in the expression screen is marked
(asterisk).
The human SARA of Tables 1 and 2, identified as described in Example 2,
regulates the subcellular localization of Smad2 and Smad3 and recruits these
Smads into distinct subcellular domains. This SARA also interacts with TGF~i
receptors and TGF(3 signaling induces dissociation of Smad2 or Smad3 from the
SARA protein with concomitant formation of Smad2/Smad4 complexes and
nuclear translocation.
Table 10 shows alignment of the amino acid sequences of the FYVE
domains from hSARAI, XSARA1, KIAA0305, FGD1, Hrs-1, Hrs-2 and EEA1.
Identical residues (dark grey) and conservative changes (light grey) are
marked.
A consensus sequence (bottom) was derived from positions in which at least 6
out of 7 residues were conserved or when proteins contained one of only two
alternate residues.
The regulation of the subcelluiar localization of components of signaling
pathways can be key determinants in the effective initiation and maintenance
of
signaling cascades. Targeting of signal transduction proteins to specific
subcellular regions is highly regulated, often through specific interactions
with
scaffolding or anchoring proteins (Faux and Scott, 1996; Pawson and Scott,
1997). Scaffolding proteins have been defined as proteins that bind to
multiple
kinases to coordinate the assembly of a cascade, while anchoring proteins are
tethered to specific subcellular regions in the cell and can act to bring
together
components of a pathway. Regulating location of signaling components can
thus coordinate the activity of a signaling network, maintain signaling
specificity
or facilitate activation of a pathway by localizing kinases together with
their
downstream substrates.
As described herein, a recombinantly produced human SARA protein
bound directly and specifically to unphosphorylated Smad2 and Smad3. In
addition, receptor-dependent phosphorylation induced Smad2 to dissociate from
SARA, bind to Smad4 and translocate to the nucleus. Thus, the hSARAI protein
functions in TGF(3 signaling upstream of Smad activation to recruit Smad2 to
the
CA 02335346 2001-O1-18
WO 00/05360 PCTlCA99/00656
19
TGF~3 receptor by mediating the specific subcellular localization of Smad and
by
associating with the TGF~i receptor complex. Furthermore, inducing
mislocalization of Smad2 by expressing a mutant of the hSARAI protein blocks
TGF(3-dependent transcriptional responses, indicating an essential role for
SARA-
mediated localization of Smads in signaling. Together, these results identify
the
cloned hSARAI protein as a novel component of the TGF~i pathway that
functions to anchor Smad2 to specific subcellular sites for activation by the
TGF~i
receptor kinase.
In vitro, receptor-regulated Smads are recognized by the receptor kinases
and are phosphorylated on the C-terminal SSXS motif (Abdollah et al., 1997;
Kretzschmar et al., 1997; Macias-Silva et al., 1996; Souchelnytskyi et al.,
1997).
This phosphorylation is similar to receptor-dependent phosphorylation in
mammalian cells, suggesting that SARA is not absolutely required for
recognition
of Smads by the receptor complex. In intact cells, however, receptor-regulated
Smads are cytosolic proteins that require activation by transmembrane
serine/threonine kinase receptors. Consequently, Smads may require
recruitment by SARA to interact with TGF~i superfamiiy receptors. Domains in
which SARA is found correspond to regions where TGF~i receptors are also
localised. TGF~i receptors display regionalized localization and hSARAI
recruits
Smad2 to these domains. The identity of these intracellular domains is
unclear.
However, they contain receptors and recent evidence has shown that FYVE
finger domains interact with membranes, so it is reasonable to suggest that
these
domains represent membrane vesicles. Thus, clustering of the TGF(3 receptor,
as
previously described by Henis et al. (1994), may function to direct the
receptor
to hSARAI and the Smad2 substrate. This activity may be most critical in vivo,
where serlthr kinase receptors are often found in low numbers and only a small
proportion need to be activated for biological responses (Dyson and Gurdoh,
1998). This activity is likely to be most critical in vivo, where ser/thr
kinase
receptors are often found in low numbers and only a small proportion need to
be
activated for biological responses (Dyson and Gurdon, 1998). This may impose
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
on the pathway a stringent requirement for SARA to anchor Smads in these sites
for receptor interaction.
The colocalization and association of hSARAI with the TGF~i receptor
defines a role for hSARAI in recruiting Smad2 to the receptor kinase.
5 Furthermore, deletion of the FYVE domain interferes with receptor binding,
prevents the correct localization of hSARAI/Smad2 and blocks TGF~i signaling
_ (see Example 8 below), suggesting that this is an important function in the
pathway. Interestingly, the binding of the hSARAI protein identified in
Example
2 to the receptor was enhanced upon Smad2 expression and, on its own, SARA
10 may interact inefficiently with the receptor. However, within the
hSARAI/Smad
complex, Smad2 might help drive association with the receptor through its
recognition of the catalytic region of the kinase domain. Consistent with
this,
cooperation requires a kinase deficient type I receptor which also traps the
Smad2 substrate (Macias-Silva et al., 1996). Thus, Smad2 may bind to the
15 catalytic pocket of the type 1 kinase domain while hSARAI, which is not a
substrate of the kinase, may interact with regions outside of the domain.
The human SARA protein identified in Example 2 did not interact with
any of the other Smads tested, indicating that it functions specifically in
Smad2
and Smad3 pathways (see Example 3). However, SmadS localization in 293
20 cells displayed a remarkably similar pattern to that of this SARA protein
(Nishimura et al., 1998) and similar patterns were observed for endogenous
Smad1 or 5 in the kidney epithelial cell line, IMCD-3. Thus, localization of
BMP-regulated Smads (for example, Smadl, SmadS and SmadB) may also be
regulated by a specific SARA family member.
The genes for two other SARA family member proteins were also
identified and cloned. One of these, identified in Xenopus and designated
XSARA2 (Tables 7 and 8), is related to XSARA 1, while the other one, hSARA2
(Tables 3 and 4), is a human clone, related to the hSARA 1 of Tables 1 and 2.
This second human clone has been identified in EST clone KIAA0305. A
comparison of the SBD from hSARAI with a similar region from the KIAA0305
sequence indicated that the amino terminal half of the region of the SBD was
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
21
highly divergent from the amino acid sequence encoded by KIAA0305. This
suggests that the protein encoded by KIAA0305 may mediate binding with other
as yet unidentified proteins, eg. other Smads. In contrast to the SBD, the
FYVE
domain of the KIAA0305 protein is more closely related to the hSARAI FYVE
5 domain (70% identity), suggesting that this protein may be an anchor for
other
Smad proteins that function either in the TGF(3 pathway or in other signaling
cascades, such as the BMP signal transduction pathway.
SARA is not limiting in Smad activation and TGF~i superfamily signaling
10 It was observed that elevating Smad2 levels can saturate hSARAI and
yield a diffuse distribution for Smad2. Thus, the level of the hSARAI protein
is a
key determinant in controlling Smad2 localization. As a consequence,
endogenous Smad2 may or may not display a hSARAI-like distribution,
depending on the relative expression of the two proteins. Indeed, in Mv1 Lu
15 cells, endogenous Smad2 displays a punctate pattern with some diffuse
staining
in the cytosol. While not meaning to limit the invention to a particular
mechanism, the data are consistent with the view that once signaling has
commenced, Smad2 dissociates from hSARAI, binds to Smad4 and translocates
to the nucleus, freeing hSARAI to recruit additional Smad2 from the cytosolic
20 reservoir. This would provide a mechanism to allow quantitative activation
of
Smads in the presence of high levels of TGF~3 signaling.
By functioning to recruit Smad2 to the TGF(3 receptor, hSARAI is located
in an important regulatory position in the pathway. Thus, control of hSARAI
localization or protein levels, or its interaction with Smad2, could modulate
25 TGF~i signaling. Further, disruption of normal hSARAI function could
potentially be involved in loss of TGF~ responsiveness that is a common
feature
during tumour progression.
Modular Domains in SARA
30 The function of hSARAI in TGF~i signaling is mediated by three
independent domains, the Smad binding domain (SBD) that mediates specific
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
22
interaction with Smad2 and Smad3, the FYVE domain that targets
hSARAI/Smad2 to specific subcellular sites and the carboxy terminus which
mediates association with the TGF~i receptor. The Xenopus and mouse
forkhead-containing DNA binding proteins, FAST1 and FAST2, bind specifically
S to Smad2 and Smad3 and like hSARAI, interact with the MH2 domains (Chen et
al., 1996; Chen et al., 1997a; Labbe et al., 1998; Liu et al., 1997a).
Comparison of the SBD from this SARA with the Smad Interaction Domain (SID)
from these FAST proteins revealed no regions of obvious similarity. However,
since hSARAI acts upstream and FAST downstream of Smad activation, these
proteins may employ structurally unrelated domains to distinguish unactivated
versus activated forms of Smad2. Thus, the SBD of this SARA protein
preferentially binds unphosphorylated monomeric Smad2 while the SID from
FAST must bind phosphoryiated Smad2 in heteromeric complexes with Smad4.
By analogy, the SBD of other SARA family members may bind the
15 unphosphorylated monomeric species of other Smads that mediate signal
transduction in other pathways (eg. Smads 1, 5 or 8 in the BMP signal
transduction pathway).
In hSARAI, the FYVE domain functions independently of the SBD, to
mediate the subcellular targetting of the protein. The FYVE-finger motif has
now
been identified in at least 30 proteins from diverse species, such as FGD1,
Hrs-1
and 2, and EEA1 (Gaullier et al., 1998; V~/iedemann and Cockcroft, 1998).
Recent advances have demonstrated that FYVE finger motifs from a variety of
divergent proteins have a conserved function and bind phosphatidyl inositol-3-
phosphate (Ptdlns(3)P) with high specificity (Burd et al., (1998); Patki
(1998);
Gaullier (1998)). Through this interaction, the FYVE finger can mediate
protein
interactions with phospholipid bilayers. However, Ptdlns(3)P is present
ubiquitously on cell membranes and in the case of EEA1, further protein-
protein
interactions with RabS-GTP are required in addition to the FYVE domain to
target the protein to the correct membranes (Simonsen et al., 1998). Given
that
Ptdlns(3)P binding by FYVE fingers is conserved in yeast and mammals, it is
likely that the FYVE finger of hSARAI similarly mediates interaction with the
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
23
membrane. Furthermore, it is possible that additional protein-protein
interactions may be required to direct hSARAI to regions that contain the
TGF~3
receptors. The carboxy terminus of hSARAI, which is required for efficient
interaction with the TGF(3 receptor, may function in this capacity.
Together, these data define discrete domains in SARA that fulfill specific
aspects of SARA function in TGF~i superfamily signaling. ~Nithout being
limited
to any particular mechanism, a possible model of the interaction of SARA with
a
receptor regulated Smad in TGF~3 superfamily signaling, as exemplified by
hSARAI and its interactions with Smad2 in TGF~3 signaling, is shown
diagrammatically in Figure 18. The FYVE domain likely functions to direct SARA
to the membrane, perhaps through interactions with Ptlns(3)P. It thus fulfills
an
important role in recruiting hSARAI to specific subcellular domains that have
been shown also to contain the TGF(3 receptor. The SBD in turn functions to
bind unactivated Smad2, thus recruiting the receptor substrate to this
subcellular
15 region. Once localized to this region, the C-terminal domain of hSARAI
functions with Smad2 bound to the SBD to promote interaction with the receptor
complex. These three domains thus function cooperatively to recruit Smad2 to
the TGF~3 receptor.
Additional Roles for SARA
Controlling the localization of kinases and their substrates may allow not
only for efficient recognition and phosphorylation but may also function to
maintain specificity and suppress crosstalk between signaling pathways. Thus,
by controlling Smad localization, a SARA family member protein could
additionally function to maintain the highly specific regulation of Smad
phosphorylation by ser/thr kinase receptors that is observed in vivo and could
prevent promiscuous phosphorylation by other kinases in the cell. Furthermore,
through its interactions with a particular receptor, a SARA protein might
function
to control the activity or turnover of the receptor complex. Alternatively,
SARA
may also fulfill scaffolding functions to coordinate the receptor-dependent
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
24
activation of Smads with other as yet unidentified components of a signaling
pathway.
Nucleic Acids
5 In accordance with one series of embodiments, the present invention
provides isolated nucleic acids corresponding to, or related to, the human and
Xenopus SARA nucleic acid sequences disclosed herein. In addition to the
SARA nucleotide sequences disclosed herein, one of ordinary skill in the art
is
now enabled to identify and isolate homologues of the SARA genes described
10 herein. One of ordinary skill in the art may screen preparations of genomic
or
cDNA from other species using probes or PCR primers derived from nucleotide
sequences disclosed herein. In accordance with a further embodiment, the
invention provides isolated nucleic acids of at least 10 consecutive
nucleotides,
preferably 15 consecutive nucleotides, more preferably 20 consecutive
15 nucleotides of Sequences ID N0:1, Sequence ID N0:3, Sequence ID N0:5 and
Sequence ID N0:7, up to the complete sequences. Short stretches of nucleotide
sequence are useful as probes or primers useful for identification or
amplification
of the nucleic acids of the invention or for encoding fragments, functional
domains or antigenic determinants of SARA proteins.
20 The invention also includes polynucleotides which are complementary to
the disclosed sequences, poiynucleotides which hybridise to these sequences at
high stringency and degeneracy equivalents of these sequences.
Proteins
25 SARA proteins may be produced by culturing a host cell transformed with
a DNA sequence encoding a selected SARA protein. The DNA sequence is
operatively linked to an expression control sequence in a recombinant vector
so
that the protein may be expressed.
Host cells which may be transfected with the vectors of the invention
30 may be selected from the group consisting of E. toll, Pseudomonas, Bacillus
CA 02335346 2001-O1-18
WO 00!05360 PCT/CA99/00656
25
subtillus, or other bacilli, yeasts, fungi, insect cells or mammalian cells
including
human cells.
For transformation of a mammalian cell for expression of a SARA protein,
the vector may be delivered to the cells by a suitable vehicle. Such vehicles
including vaccinia virus, adenovirus, retrovirus, Herpes simplex virus and
other
vector systems known to those of skill in the art.
A SARA protein may also be recombinantly expressed as a fusion protein.
For example, the SARA cDNA sequence is inserted into a vector which contains
a nucleotide sequence encoding another peptide (e.g. GST-glutathione succinyl
transferase). The fusion protein is expressed and recovered from prokaryotic
(e.g. bacterial or baculovirus) or eukaryotic cells. The fusion protein can
then be
purified by affinity chromatography based upon the fusion vector sequence and
the SARA protein obtained by enzymatic cleavage of the fusion protein.
The protein may also be produced by conventional chemical synthetic
methods, as understood by those skilled in the art.
SARA proteins may also be isolated from cells or tissues, including
mammalian cells or tissues, in which the protein is normally expressed.
The protein may be purified by conventional purification methods known
to those in the art, such as chromatography methods, high performance liquid
chromatography methods or precipitation.
For example, anti-SARA antibodies may be used to isolate SARA protein
which is then purified by standard methods.
Antibodies
The provision of the polynucleotide and amino acid sequences of SARA
proteins provides for the production of antibodies which bind selectively to a
SARA protein or to fragments thereof. The term "antibodies" includes
poiyclonal antibodies, monoclonal antibodies, single chain antibodies and
fragments thereof such as Fab fragments.
30 In order to prepare polyclonal antibodies, fusion proteins containing
defined portions or all of a SARA protein can be synthesized in bacteria by
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
26
expression of the corresponding DNA sequences, as described above. Fusion
proteins are commonly used as a source of antigen for producing antibodies.
Alternatively, the protein may be isolated and purified from the recombinant
expression culture and used as source of antigen. Either the entire protein or
fragments thereof can be used as a source of antigen to produce antibodies.
The purified protein is mixed with Freund's adjuvant and injected into
rabbits or other appropriate laboratory animals. Following booster injections
at
weekly intervals, the animals are then bled and the serum isolated. The serum
may be used directly or purified by various methods including affinity
chromatography to give polyclonal antibodies.
Alternatively, synthetic peptides can be made corresponding to antigenic
portions of a SARA protein and these may be used to inoculate the animals.
In a further embodiment, monoclonal anti-SARA antibodies may be
produced by methods well known in the art. Briefly, the purified protein or
fragment thereof is injected in Freund's adjuvant into mice over a suitable
period
of time, spleen cells are harvested and these are fused with a permanently
growing myeloma partner and the resultant hybridomas are screened to identify
cells producing the desired antibody. Suitable methods for antibody
preparation
may be found in standard texts such as Antibody Engineering, 2d. edition,
Barreback, ED., Oxford University Press, (1995).
Trans~enic animals
in accordance with a further embodiment, the invention provides for the
production of transgenic non-human animals which afford models for further
study of the SARA family of proteins and also provide tools for the screening
of
the candidate compounds as therapeutics.
Animal species which are suitable for use include rats, mice, hamsters,
guinea pigs, rabbits, dogs, cats, goats, sheep, pigs and non-human primates.
in accordance with one embodiment, a transgenic animal may be
prepared carrying a heterologous SARA gene by inserting the gene into a germ
line or stem cell using standard technique of oocyte microinjection, or
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
27
transfection or microinjection into embryonic stem cells. The techniques of
generating transgenic animals are now well known and fully described in the
literature. For example, a laboratory manual in the manipulation of the mouse
embryo describes standard laboratory techniques for the production of
transgenic mice (Hogan et al. (1986), Manipulating the Mouse Embryo, Cold
Spring Harbour Laboratory Press, Cold Spring Harbour, New York).
In accordance with a further embodiment, the invention enables the
inactivation or replacement of an endogenous SARA gene in an animal by
homologous recombination. Such techniques are also fully described in the
literature. Such techniques produce "knock-out" animals, with an inactivated
gene, or "knock-in" animals, with a replaced gene.
EXAMPLES
The examples are described for the purposes of illustration and are not
intended to Limit in any way the scope of the invention.
Methods of molecular genetics, protein and peptide biochemistry and
immunology referred to but not explicitly described in this disclosure and
examples are reported in the scientific literature and are well known to those
ski I led i n the art.
Example 1: Methods
Isolation of Xenopus and human SARA
To prepare a probe for library screening, the MH2 domain of Smad2
(amino acids 241-467) was subcloned into a modified pGEX4T-1 vector
containing the protein kinase A recognition site derived from pGEX2TK
(Pharmacia). This bacterial fusion protein was purified, labelled with
[3zP]YATP
and used as probe to screen a ,ZAP II Xenopus dorsal lip library as described
(Chen and Sudol, 1995). A screen of 1 x 10~ plaques yielded four phage which
represented repeated isolates of the same clone. This partial cDNA contained a
2.i kb open reading frame and 1 kb of 3' untranslated region (UTR). A full
length clone was obtained by a combination of rescreening of the same dorsal
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
28
lip library using a 670 base pair EcoRl/Hpal fragment at the 5' end of this
clone
and by 5' RACE (Gibco/BRU using stage 10 Xenopus RNA.
To obtain a human homolog of Xenopus SARA, cDNA was synthesized
from randomly primed total RNA isolated from HepG2 cells. This cDNA was
subjected to polymerase chain reaction (PCR) using degenerate primers as
described previously (Attisano et al., 1992). The 5' and 3' primers, designed
to
encode the zinc-finger motif, correspond to
GC(A/C/GITnCC(A/C/G/T)AA(CmTG(C/TATGAA(A/C/G/T)TG(C/T) and
(A/G)CA(A/G)TA(C/T)TC(A/CJG/T)GC(A/C/G/T)GG(A/G)TT(A/G)TT, respectively.
A 150 base pair PCR product was sequenced and then used as probe for
screening a 7LZAP human fetal brain cDNA library (Stratagene). Eight positive
plaques were obtained, two of which contained an overlap of approximately 1 kb
and covered the entire open reading frame. The sequence of the 5' UTR was
confirmed by sequencing of an expressed sequence tag database clone (clone ID
260739).
Construction of Plasmids
For mammalian expression constructs of SARA, the open reading frame of
hSARA was amplified by PCR and was subcloned into pCMVS in frame with an
amino-terminal Flag or Myc tag (Hoodless et al., 1996). The deletion mutants
of
pCMVS-Flag-hSara0893-1323, 0346-132, 0893-1323, and 0346-1323 were
constructed by deletion of EcoRV-Hindlll, Xbal-Hindlll, Sall-EcoRV, and Sall-
Xbal
fragments, respectively. PCMVS-Flag-hSara01-594 and 01-686 were obtained
by partial digestion with Asp718/Sall and for pCMVS-Flag-hSARA X665-1323 a
Asp718/Hindlll partial digest was used. PCMVS-Flag-hSARA0596-704 was
constructed by deleting Asp718 fragment. The other hSARA mutants were
constructed by PCR using appropriate primers. PCMVSB-Myc-Smad3 and Myc-
Smad6, pGEX4T-1-Smad2/MH1 (amino acids 1-181), pGEX4T-1-Smad2/linker
(amino acids 186-273), pGEX4T-1-Smad2/MH2 (amino acids 241-467) and
pGEX4T-1-h SARA (amino acids 665-750) were constructed by PCR.
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
In Vitro Protein Interactions
29
In vitro transcription/translation reactions were performed using the TNT
coupled reticulocyte lysate system (Promega) following the manufacturer's
instructions using T3 RNA poiymerase. Translation was carried out in the
presence of [35S]-methionine and labelled proteins were incubated with
purified
GST fusion proteins in TNTE buffer with 10% glycerol for 2 hours at
4°C and
then washed five times with the same buffer. Bound protein was separated by
SDS-PAGE and visualized by autoradiography.
Immunoprecipitation and Immunoblotting
COS-1 cells transfected with LipofectAMINE (GIBCO BRL) were lysed
with lysis buffer (Wrana et al., 1994) and subjected to immunoprecipitation
with
either anti-Flag M2 (IBI, Eastern Kodak) or anti-Myc (9E10) monoclonal
antibody
followed by adsorption to protein-G sepharose. Precipitates were separated by
SDS-PAGE, transferred to nitrocellulose membranes and immunoblotted as
described previously (Hoodless et al., 1996).
Affinity-Labelling
LipofectAMINE transfected COS-1 cells were incubated with 200 pM
['zSIJTGF(3 in media containing 0.2% bovine fetal serum at 37°C for 30
minutes
and receptors were cross-linked to ligand with DSS as described previously
(Macias-Silva et al., 1996). Cell fysates were immunoprecipitated with anti-
Flag
antibody and receptors visualized by SDS-PAGE and autoradiography. in some
cases, cross-linked ['251]TGF(3 was determined by gamma counting.
Subcellular Localization by Immunofluorescent Confocal Microscopy
Mv1 Lu cells, plated on gelatin-coated Permanox chamber slides (Nunc),
were transfected by the calcium phosphate-DNA precipitation method. Fixation,
permeabilisation and reaction with the primary and secondary antibodies were
described previously (Hoodless et al., 1996). Monoclonal anti-Flag antibodies
were visualized by FITC-conjugated goat anti-mouse IgG (Jackson Laboratories)
CA 02335346 2001-O1-18
yV0 00/05360 PCT/CA99/00656
30
and polyclonal Myc antibody (A14, Santa Cruz) was visualized with Texas-Red-
conjugated goat anti-rabbit IgG (Jackson Laboratories). Immunofluorescence was
analyzed on a Leica confocal microscope.
Transcriptional Response Assay
Mv1 Lu cells were transiently transfected with the reporter plasmid, p3TP-
lux (Wrana et al., 1992), CMV-(3gal and selected constructs using calcium
phosphate transfection. Twenty-four hours after transfection, cells were
incubated overnight with or without SO pM TGF[i. Luciferase activity was
measured using the luciferase assay system (Promega) in a Berthold Lumat LB
9501 luminometer and was normalized to [i-galactosidase activity.
Examale 2 - Identification of SARA family members
The MH2 domain of Smad2 was fused to glutathione-S-transferase (GST)
that included a kinase recognition site for protein kinase A (PKA). The
bacterially-expressed fusion protein was labelled to high specific activity
using
PKA (Chen and Sudol, 1995), and then used to screen a 7~ZAP11 expression
library prepared from the dorsal blastopore iip of Xenopus. From this screen,
four clones were identified, all of which presented a repeated isolate of a
partial
cDNA clone with no similarity to sequences in the GenBank database. To
confirm that the product encoded by this clone interacted with Smad2, an in
vitro transcription/transiation system was used to produce [35SJmethionine-
labelled protein. Translation of the cDNA yielded a protein product of
approximately 80 kDa which corresponded in size to the longest open reading
25 frame (ORF) identified in the sequence. Incubation of this product with
bacterially-produced GST-Smad2(MH2) resulted in efficient binding of the
translated product to the fusion protein (data not shown). Interaction with
full
length Smad2 was also observed, whereas binding to bacterially-expressed
Smad 1 or Smad4 was not.
To isolate a full length cDNA, the partial clone identified in the
interaction screen was used as a probe to rescreen the same blastopore lip
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
31
library. Since the resulting clones lacked the 5' end, 5' RACE was conducted
to
obtain the entire coding sequence. Analysis of the complete cDNA sequence
(Table 5) revealed a long open reading frame that was contiguous with that of
the partial clone. The predicted protein, XSARA1, is 1235 amino acids long
with
5 an estimated molecular mass of 135 kDa (Table 6). Analysis of the full
length
cDNA sequence (Table 9) revealed a region in the middle portion of the
predicted protein that had similarity to a double zinc finger domain (recently
renamed the FYVE domain; Mu et al., 1995). The FYVE domain has been
identified in a number of unrelated signaling molecules that include FGD1, a
10 putative guanine exchange factor for Rho/Rac that is mutated in
faciogenital
dyspiasia (Pasteris et al., 1994), the HGF receptor substrate Hrs-1 and its
homolog Hrs-2 (Bean et al., 1997; Komada and Kitamura, 1995), EEA1, a protein
involved in formation of the early endosome (Mu et al., 1995) and the yeast
proteins FAB1, VPS27 and VAC1 (Piper et al., 1995; Weisman and Wickner,
1 S 1992; Yamamoto et al., 1995). Comparison of the FYVE domains from the
vertebrate proteins with that from SARA revealed extensive conservation of
residues throughout the domain (Table 10). Thus, SARA contains a FYVE
domain that may fulfill important functions in diverse proteins.
To investigate the role of SARA in TGF~3 superfamily signaling in
20 mammalian cells, a human homologue was identified. Using a carboxy-terminal
portion of XSARA1, a human library was screened and a protein was identified
that was distantly related to Xenopus SARA (34°/° identity) and
which was also
sequenced as an EST (KIAA0305). However, no homologs closer to XSARA
were identified. Thus, degenerate oligonucleotide primers were designed
25 encoding amino acids in XSARA1 (Table 9) and HepG2 RNA was used as
template for degenerate PCR. A related sequence was identified and this
partial
cDNA was used to screen a human brain cDNA library. Four overlapping
clones, encoding a long open reading frame were identified and a search of the
EST database with this sequence led to the identification of additional
30 overlapping cDNA clones from libraries derived from T cells, uterus,
endothelial
cells and melanocytes. Analysis of the contiguous sequence revealed a long
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
32
open reading frame that had a consensus start codon preceded by stop codons in
all three reading frames (Table 1). Comparison of the predicted protein hSARAI
(Table 2), from this cDNA with XSARA1 (Table 9) revealed an overall identity
of
62%, with a divergent 558 residue amino terminal domain (35°/°
identity)
followed by a closely related carboxy terminus (85% identity).
Example 3 - hSARA interacts specificaliy with Smad2 and Smad3
To characterize the interaction of hSARA with Smads, the full length
protein was translated in vitro and tested for binding to bacterially-
expressed
10 Smad fusion proteins. Similar to the Xenopus clone, hSARAI bound
specifically
to full length Smad2, but not Smad1 or Smad4 (Figure 1). In addition, full
length
Smad3, which is highly related to Smad2, also interacted with hSARAI. To
define the domains of Smad2 that bound hSARA, in bacteria various fragments of
Smad2 corresponding to the MH1 domain, linker region and MH2 domain were
15 expressed in bacteria. Similar to the Xenopus clone, hSARA interacted
efficiently
with fusion proteins that comprised the MH2 domain, while no association was
detected between hSARA and either the MH1 or non-conserved linker domains
(Figure 1 ). Thus, hSARAI specifically interacts with Smad2 through the MH2
domain.
20 To confirm that hSARA also bound to Smads in mammalian cells, a Flag
epitope tag was introduced at the amino terminus of the protein to create Flag-
SARA. Transient expression of Flag- SARA in COS cells yielded a protein of the
predicted molecular weight for SARA (Figure 2) that was not present in
untransfected cells (data not shown). To investigate the interaction of SARA
with
25 Smads, Flag- SARA was expressed in COS cells together with Myc-tagged
versions of Smads 1, 2, 3, 4, 6 and 7. Cell lysates were subjected to anti-
Flag
immunoprecipitation followed by immunoblotting with anti-Myc antibodies. In
other immunoprecipitates of cells expressing either Smad2 or Smad3, efficient
coprecipitation of either Smad with Flag- hSARAI was observed (Figure 2). In
30 contrast, none of the other Smads coprecipitated with hSARAI. Specific
binding
of this SARA family member to both Smad2 and Smad3 is consistent with the
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
33
observation that these two proteins possess very closely related MH2 domains
(97°/° identity) and are both activated by TGF~i or activin type
I receptors (Liu et
a., 1997b; Macias-Silva et al., 1996; Nakao et al., 1997a). Together, these
results demonstrate that this SARA family member is a specific partner for
receptor-regulated Smads of the TGF~3/activin signaling pathway.
Example 4- Phosphorytation of Smad2 induces dissociation from SARA
Previous findings have shown that activation of TGF(3 signaling results in
phosphorylation of Smad2 or Smad3 by type I receptors on C-terminal serine
residues (Liu et al., 1997b; Macias-Silva et al., 1996). A constitutively
active
TGF~i type I receptor was prepared by substituting a threonine in the GS
domain
with an aspartate residue (Wiener et al., 1995). This activated type I
receptor
induces TGF(3 signaling in the absence of type II receptors and ligand and
regulates the phosphorylation and activation of Smad proteins in a manner
similar to ligand (Macias-Siiva et al., 1996; Wiener et al., 1995). COS cells
were
transfected with combinations of Smad2, hSARAI or both in the presence or
absence of activated T~iRI. Cells were then metabolically labelled with
[3~P]phosphate and phosphorylation of either hSARAI or Smad2 was assessed in
immunoprecipitates. Analysis of SARA phosphorylation revealed that the protein
was banally phosphorylated and the coexpression of the activated type I
receptor
did not appreciably affect the overall phosphorylation (Figure 3). In
contrast,
analysis of Smad2 immunoprecipitated from total cell lysates showed that the
activated type I receptor induced strong phosphorylation of the protein as
described previously (Macias-Silva et al., 1996). These results suggest that
SARA
is not phosphorylated in response to TGF(3 signaling.
The phosphorylation state of Smad2 that coprecipitated with hSARAI was
examined. Interestingly, unlike the strong induction of Smad2 phosphorylation
in the total cellular pool, phosphorylation of Smad2 associated with hSARAI
was
not enhanced, but rather appeared to decrease in the presence of TGF~i
signaling
(Figure 3). This suggested that receptor-dependent phosphorylation of Smad2
might induce dissociation from hSARAI . To examine this directly, the
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
34
interaction of hSARAI with wild type Smad 2 or a mutant version lacking the C-
terminal phosphorylation sites (Smad2(2SA)) was analysed. in the absence of
TGF~i signaling, association of hSARAI with either Smad2 or Smad2(2SA) was
comparable (Figure 4). In contrast, in cells coexpressing the activated
receptor, a
significant decrease in the interaction of wild type Smad2 with hSARAI was
observed. However, hSARAI/Smad2(2SA) complexes were not reduced by the
activated receptor. Together, these results suggest that hSARAI is not
phosphorylated in response to TGF~3 signaling and that it preferentially
interacts
with the unphosphorylated form of Smad2.
Example 5 - SARA and Smad4 form mutually exclusive complexes with Smad2
Phosphorylation of Smad2 induces its interaction with Smad4 (Lagna et
al., 1996; Zhang et al., 1997). hSARAI/Smad2 complexes in COS cells
coexpressing Smad4 were assessed. In unstimulated cells, the level of
hSARAI/Smad2 complex formation was comparable either in the presence or
absence of Smad4 (Figure 5, lanes 3 and 6). However, upon activation of TGF(3
signaling, dissociation of Smad2 from hSARAI was significantly enhanced by
coexpression of Smad4 (Figure 5, lanes 4 and 7). These results indicated that
phosphorylated Smad2 might preferentially interact with Smad4 rather than
hSARAI and suggested that Smad2 might form mutually exclusive complexes
with either Smad4 or hSARAI . The formation of Smad2/Smad4 and
Smad2/hSARA4 complexes in the same transfectants was then examined. Cell
lysates were subjected to immunoprecipitation with anti-Flag antibodies
directed
towards tagged Smad2 and then immunoblotted for the presence of Smad4 and
hSARAI . Consistent with previous findings (Lagna et al., 1996; Zhang et al.,
1997), interaction of Smad4 with Smad2 was strongly stimulated by the
activated
type I receptor (Figure 6, lane 3 and 4). Concomitant with the formation of
Smad2/Smad4 complexes, the interaction of Smad2 with hSARAI was disrupted
by activation of signaling (Figure 6, lanes 6 and 7). Thus, complexes of
Smad2/hSARAI and Smad2/Smad4 are mutually exclusive, supporting the notion
that Smad4 may compete for Smad2 to enhance dissociation of hSARAI/Smad2
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
complexes. Together these results demonstrate that during TGF~i signaling,
hSARAI/Smad2 complexes are transient and phosphorylation of Smad2 induces
dissociation and formation of heteromeric complexes with Smad4.
5 Example 6- hSARAI regulates the subcellular localization of Smad2
The studies described above suggest that SARA functions upstream in the
pathway and might control the subcellular localization of Smad2. To test this,
an investigation was done to determine whether coexpression of hSARAI might
alter the localization of Smad2 in the TGF(3-responsive epithelial cell line,
10 Mv1 Lu, using confocal microscopy. Mv1 Lu cells were used rather than COS
since the Myc antibodies crossreacted with endogenous proteins in the COS and
obscured nuclear staining of tagged proteins. In cells expressing hSARAI
alone,
the protein displayed a punctate staining pattern that was present throughout
the
cytosolic compartment and was excluded from the nucleus (Figure 7A). This
15 localization of hSARAI was in contrast to the diffuse staining typically
observed
for Smad2 in cells overexpressing the protein (Figure 7B). Cells transiently
transfected with both hSARAI and Smad2 were examined. In these cells, the
distribution of hSARAI was indistinguishable from cells transfected with
hSARAI
alone (Figure 7D, left photo). In contrast, the localization of Smad2 in the
20 presence of hSARAI displayed a dramatic shift to a punctate pattern
(compare
Figure 7B to 7D, centre photos). Moreover, analysis of these immunofluorescent
staining patterns by confocal microscopy revealed that hSARAI and Smad2
precisely colocalized in the cytosol (yellow stain, Figure 7D, right photo).
Interestingly, expression of Smad2 at much higher levels than hSARAI reverted
25 the distribution of Smad2 to that observed in cells transfected with Smad2
alone
(data not shown). This supports the notion that elevating the amount of Smad2
can saturate hSARAI and yield a diffuse distribution of Smad2 throughout the
cell.
Studies were conducted to determine whether activation of TGF~i
30 signaling induces nuclear translocation of Smad2 in the presence of hSARAI
. As
shown in Figure 7, the localization of hSARAI in the cytosolic compartment
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
36
looked similar in the presence or absence of the constitutively active TGF~i
type I
receptor (compare Figure 7D and E, left photos). However, TGFJ3 signaling
caused a significant proportion of Smad2 to translocate to the nucleus (Figure
7E,
centre photo) and this correlated with a shift to an orangy-red colour in the
cytosolic colocalization stain (Figure 7E, right photo). Thus activation of
TGF(3
signaling induces Smad2 to dissociate from hSARAI and translocate to the
nucleus.
To confirm that the punctate localization of overexpressed SARA reflected
that of the endogenous protein, the localization of endogenous SARA and Smad2
10 was examined in Mv1 Lu cells. Analysis of the distribution of endogenous
hSARAI using affinity-purified rabbit anti-hSARAI antibodies revealed a
punctate distribution that was similar to the pattern observed for transiently
transfected, epitope-tagged hSARAI (Figure 7F, left photo). This staining was
specific, since cells stained with preimmune antisera, or purified antibody
15 blocked with the hSARAI antigen, revealed no detectable staining in the
cytosol,
although some weak background staining was observed in the nucleus (data not
shown). Examination of endogenous Smad2 distribution in the same cell using
goat anti-Smad2 antibodies revealed a punctate distribution for Smad2 (Figure
7F, centre photo) as published previously (Janknecht et al., 1998).
Furthermore,
20 analysis of hSARAI and Smad2 together revealed extensive colocaiization of
the
two proteins (Figure 7F, right photo). Colocalization was not complete and may
reflect differences in the stoichiometry of hSARAI versus Smad2 protein levels
as
suggested above, or the presence of additional regulatory mechanisms in the
cell
that control interaction of the endogenous proteins.
25 Taken together with the biochemical analysis, these results indicate that
hSARAI functions to anchor or recruit Smad2 to specific subcellular regions
prior to activation by TGFJ3 signaling.
Example 7 - hSARAI co-localises with TJ3R11
30 The positioning of hSARAI upstream of Smad2 activation suggested to us
that hSARAI might recruit Smad2 to specific subcellular domains for
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
37
phosphorylation and activation by the receptor. interestingly, previous
studies
on the TGF~i receptor demonstrated clustering of the receptor complex into
punctate domains that resembled those displayed by hSARAI (Henis et al.,
1994). To test whether hSARAI might colocalize with TGF~3 receptors, the
subcellular localization of hSARA1 and TGF~i Mv1 Lu receptors was investigated
in Mv1 Lu cells. Endogenous TGF(3 receptors could not be detected, likely due
to the low numbers of TGF(3 receptors present on these cells and the even
fewer
number that are activated in the presence of ligand. The localization of
hSARAI
in Mv1 Lu cells cotransfected with T~iRll and treated with TGFj3 was therefore
10 examined. In the absence of hSARAI, T(3R11 displayed a punctate staining
pattern similar to the hSARAI pattern (Figure 8A, panels i and ii,
respectively), as
observed previously in COS cells. Furthermore, in cells coexpressing hSARAI
and TGF~3 receptors, extensive colocalization of hSARAI and T~3R11 was
observed (Figure 8A, panel iii). This colocalization was not complete. This
may
15 be due to a restricted distribution of hSARAI in only a subset of the
intracellular
compartments normally occupied by transmembrane receptors, which include
the endoplasmic reticulum, Golgi and endocytic pathways. Thus, hSARAI and
the TGF~3 receptors colocalize to common subcellufar domains.
The colocalization of hSARAI and the TGF~i receptors suggested the
20 possiblity that hSARAI may interact with the TGF(3 receptor. To test this,
a
strategy was utilised similar to that employed to characterize the interaction
of
Smad2 with the TGF(3 receptor (Macias-Silva et al., 1996). Briefly, COS cells
were cotransfected with TGF~3 receptors in the presence of hSARAI and were
affinity-labelled using ['zsl]TGF~i. hSARAI was then immunoprecipitated from
25 the cell lysates and coprecipitating receptor complexes were resolved by
SDS-
PAGE and visualized by autoradiography or were quantitated using a gamma
counter. Analysis of cells expressing wild type receptors type II and type I;
revealed that receptor complexes coprecipitated with hSARAI (Figure SB, lane
3). Furthermore, in the presence of kinase deficient type I receptor, there
was a
30 small increase in binding of hSARAI to the receptor (Figure 8B, lane 2).
This is
in contrast to Smad2, which only interacts with TGF[3 receptor complexes that
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
38
contain kinase deficient type I receptors (Macias-Silva et al., 1996). These
data
suggest that hSARAI associates with the TGF~ receptor.
Next examined was whether coexpression of Smad2 might enhance the
interaction of hSARAI with TGF(3 receptors. In cells expressing wild type
receptor I, no difference in the amount of receptor complexes that
coprecipitated
with hSARAI, either in the presence or absence of Smad2, was observed (Figure
8B, compare lanes 3 and 5). In contrast, the association of hSARAI with
receptor complexes containing kinase-deficient type I receptors was enhanced
by Smad2 (Figure 8B, lane 4). This finding was consistent with the previous
demonstration that kinase-deficient type I receptors stabilize interactions of
Smad2 with the receptors. To investigate further the requirement for Smad2 in
the interaction of hSARAI with the receptor, a mutant of hSARAI, SARA(OSBD),
that removes the Smad binding domain, was tested. Analysis of wild type
hSARAI interaction with receptor complexes containing kinase-deficient T~3R1
showed that wild type hSARAI interacted with the receptor and this was
enhanced approximately two-fold by Smad2 (Figure 9A). The OSBD mutant of
hSARAI retained the capacity to associate with the receptor, although the
efficiency of interaction was slightly reduced relative to wild type hSARAI .
Importantly, unlike wild type hSARAI, binding of mutant hSARAI to the
receptor was not enhanced by coexpression of Smad2. Together, these data
suggest that hSARAI interacts with the TGF(3 receptor independently of Smad2
binding and that Smad2 cooperates to enhance the association.
To further characterize the domains in SARA that mediate binding to the
TGF(3 receptor, the interaction of a panel of SARA mutants with the TGF(3
receptor was tested. Interestingly, interaction with the TGF~3 receptor was
strongly suppressed in three mutants in which the FYVE domain was disrupted
(Figure 9B; 0594, 0664 and the internal deletion 0597-665). Since the FYVE
domain is required for the correct subcellular localization of SARA, it was
postulated that, once bound to the membrane, other regions in SARA might
contribute to the interaction with the receptor. To examine this possibility,
several carboxy-terminal truncation mutants of hSARAI were tested.
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
39
Interestingly, deletion of the C-terminus downstream of position 750
suppressed
receptor interaction, despite efficient expression of the truncated protein.
This
suggests that regions in the carboxy-terminus of SARA contribute to receptor
interaction. In these analyses, the question of whether overexpression of
Smad2
could rescue some interaction of SARA mutants with the receptor was also
explored. For both the FYVE domain mutants and the C-terminal truncation,
Smad2 expression was able to restore some interaction with the TGF~3 receptor.
It is likely that the high levels of protein and receptor expression that are
achieved in COS cells can drive some receptor interaction, even in the absence
of appropriate localization signals.
Example 8 - A modular domain in SARA mediates association with Smads
To investigate the functional importance of SARA in TGF(3 signaling, the
domains in the protein that mediate both its localization to specific
subcellular
15 regions and its interaction with Smad2 were defined. To this end, a series
of
deletion mutants of hSARAI were constructed and tested for their ability to
interact with Smad2 in COS cells by immunoprecipitation followed by
immunoblotting. As summarized in Figure 10, loss of the first 665 amino acids
of hSARAI, which included the double zinc finger/FYVE domain, did not
interfere with hSARAI binding to Smad2. However, further deletions (01-704)
completely abolished the interaction of Smad2 with hSARAI . To map the
carboxy-terminal boundary of the Smad binding domain, a number of C-terminal
truncations were also analyzed. Deletion of all residues downstream of
position
750 did not affect Smad2 interaction with hSARAI, while an additional loss of
85 amino acids (0665-1323) completely abrogated binding to Smad2. To
determine whether the region defined by this deletiona) analysis was
sufficient to
bind Smad2, the 85 amino acids referred to as the Smad Binding Domain (SBD)
were linked to GST and the fusion protein was expressed in bacteria (GST-h
SARA(665-750)). Incubation of lysates prepared from cells expressing Smad2 or
30 Smad3 with GST-SBD resulted in efficient binding of both Smads to the
fusion
protein (Figure 11A). This interaction is likely direct, since bacterially
expressed
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
40
SBD associates efficiently with bacterially-produced Smad2 (data not shown).
These studies thus define a novel domain in SARA that mediates interaction
with
Smad2 and Smad3 and which is located downstream of the FYVE domain.
The above-described analysis in COS cells showed that phosphorylation
of Smad2 by the TGF(3 receptor induced dissociation from SARA. To determine
whether this reflects an alteration in the ability of the SBD to bind
phosphorylated Smad2, the interaction of GST-SBD with Smad2 in lysates
obtained from cells expressing Smad2 alone, or Smad2 together with either wild
type or activated TGF~i type I receptor, was tested. As described previously,
10 coexpression of activated type I receptors with the appropriate receptor-
regulated Smad yields efficient phosphorylation of Smad protein. In lysates
from
cells expressing Smad2 alone or Smad2 with wild type receptors, efficient
binding of Smad2 to GST-SBD was observed. In contrast, in the presence of
activated Tj3Rl, the interaction of Smad2 with GST-SBD was strongly reduced
15 (Figure 11 B). This reduction correlated with receptor-dependent
phosphorylation, since the phosphorylation site mutant, Smad2(2SA), interacted
efficiently with GST-SBD, even in the presence of activated T(3R1 (data not
shown). These data strongly support a mechanism whereby SARA interacts with
unphosphorylated Smad2 and receptor-dependent phosphorylation induces
20 dissociation by altering the affinity of Smad2 for the SBD.
Examine 9 - The FYVE domain controls the subcellular localization of SARA
The subcellular localization of a selection of the SARA mutants was
analysed by immunofiuorescence and confocal microscopy. Analysis of
25 truncation mutants that removed the amino terminus upstream of the FYVE
domain (D1-531) yielded wild type patterns of staining (Figure 12, compare
panels i and ii). However, a further deletion (D1-664) that disrupted the FYVE
domain but did not int=-~rfere with the Smad binding domain, abolished the
wild
type staining pattern (Figure 12, panel iii). Similar studies of the C-
terminal
30 domains showed that residues downstream of the FYVE domain (0665-1323) did
not alter the localization of the mutant protein (Figure 12, panel iv), while
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
41
truncations within the FYVE domain (0596-1323) led to diffuse localization
throughout the cell (Figure 12, panel v). Of note, the 0665-1323 mutant lacked
the Smad binding domain, thereby indicating that interaction with Smad2 is not
required for proper SARA localization. To confirm that FYVE domain function
S was required for localization of SARA, a mutant with a small internal
deletion
that removes the FYVE domain 0597-664) was tested. Consistent with the other
mutants, localization of this protein was clearly disrupted (Figure 12, panel
vi).
Since none of these mutants interfered with Smad binding, the FYVE domain
appears to be required to maintain the normal localization of SARA but is not
involved in mediating interactions with Smads.
Examale 10 - SARA-mediated localization of Smad2 is necessary for TGFfi
si~nalin;g
The availability of mutants of hSARAI that interact with Smad2 but fail to
target to the appropriate subcellular sites allowed the question of whether
hSARAI-mediated localization of Smad2 was important to TGF~3 signaling to be
addressed. Whether SARA(01-594) and SARA(01-664), which bind Smad but
fail to distribute to the correct subcellular domains, would mislocalize Smad2
was examined. Coexpression of either mutant with Smad2 showed that they
20 were unable to recruit Smad2 to the normal SARA domains (Figure 13A, panels
i
and ii). As expected, SARA(01-704), which lacks a Smad binding domain, was
unable to control Smad2 localization (Figure 13A, panel iii). Whether these
mutants could cause mislocalization of Smad2 was also examined. For this,
cells were cotransfected with wild type hSARAI and Smad2 either in the
absence or presence of SARA(~1-594), SARA(01-664) or SARA(01-704). In
control transfectants, performed in the absence of mutant hSARAI, hSARAI and
Smad2 were colocalized in punctate domains as described above (Figure 13B,
panel i). However, in the presence of either SARA(O1-594) or SARA(01-664),
the localization of wild type hSARAI was normal, but the distribution of Smad2
30 was clearly disrupted and displayed a diffuse pattern (Figure 13B, panels
ii and
iii, respectively). Moreover, coexpression of SARA(~1-704), which does not
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
42
bind Smad2, resulted in Smad2 distribution that was indistinguishable from
that
of the wild type pattern (Figure 13B, panel iv). Thus, SARA(~1-594) and
SARA(~1-664) induce the mislocalization of Smad2.
Since SARA(~1-664) mislocalizes Smads and interferes with receptor
association, we investigated whether this mutant would disrupt TGF~i
signaling.
To test this, we transiently transfected the TGF(3-responsive reporter gene
3TP-
lux into Mv1 Lu cells in the presence and absence of wild type or mutant
versions of hSARAI . Expression of wild type hSARAI had no effect on TGF~i
signaling (Figure 14). In contrast, transfection of SARA(01-664) significantly
inhibited TGF(3-dependent signaling at the lowest concentration of DNA tested,
while transfection of higher doses completely abolished responsiveness of the
cells. We also tested SARA(01-704) which Packs a functional Smad binding
domain and does not alter Smad2 localization. Transfection of this mutant had
no effect on TGF(3 signaling (Figure 14). In addition to analysis of the 3TP
promoter, we examined induction of the activin response element (ARE) from
the Xenopus Mix.2 gene in HepG2 cells.
This ARE is stimulated by either TGF~i or activin signaling, which induces
assembly of a DNA binding complex that is composed of Smad2, Smad4 and a
member of the FAST family of forkhead DNA binding proteins. Since HepG2
cells do not possess endogenous FAST activity, wild type or mutants of hSARAI
were cotransfected with FAST2 and the ARE-lux reporter plasmid as described
previously (Labbe et al., 1998). Expression of either SARA(1-0594) or SARA(1-
0664), which interfere with or delete the FYVE domain, respectively, resulted
in
a strong suppression of TGF(3-dependent induction of the ARE (Figure 15).
However, none of the other mutants tested suppressed activation of this
promoter. Since none of these latter mutants disturb the localization of
hSARAI-
Smad2 complexes, these data strongly suggest that recruitment of Smad2 to the
receptor-containing subcellular domains is important for TGF/3 signaling.
Example 11 - Tissue distribution of hSARA exlaression
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
43
The 3'UTR of hSARAI and a Smad2 cDNA fragment were used to probe
a human multiple tissue Northern blot (Clontech). The results are shown in
Figure 16 - hSARAI : upper panel and Smad2: lower panel. hSARAI and Smad2
were ubiquitously expressed in the tissues examined; relatively low levels of
hSARAI were selected in liver. hSARAI and Smad2 showed a similar
expression pattern except in placenta, where proportionally more Smad2
message was observed. A single transcript of 5.0 kb is seen, corresponding to
the full length hSARAI cDNA.
SARA expression was examined in a variety of cell lines using RT-PCR
10 analysis and the gene was found to be expressed in every cell line tested.
These
included HepG2 hepatoma cells, NBFL neuroblastoma cells, SW480 colorectal
cancer cells, N 1 H 3T3 fibroblasts, P19 embryonic carcinoma cells, MC3T3
calvarial cells and Mv1 Lu lung epithelial cells (data not shown). hSARAI
appears to be a ubiquitously expressed partner for Smad2 and Smad3.
Example 12 - Interaction of endogenous hSARA1 and Smad2 in mammalian
cells
Lysates from HepG2 cells, either untreated or treated with InM TGF~i,
were immunoprecipitated with an affinity-purified, anti-hSARAI rabbit
polyclonal antibody and the immunoprecipitates were immunoblotted with a
polyclonal, anti-Smad2 antibody (Macias-Silva et al., 1998). Controls were
immunoprecipitated with pre-immune sera or N19 anti-Smad2/3 antibody. The
results are shown in Figure 17. In immunoprecipitates prepared with
preimmune antisera, no Smad2 was detectable. Anti-hSARAI
immunoprecipitates clearly showed Smad2 co-precipitating with hSARAI . TGF~3
treatment prior to lysis gave decreased association of Smad2 and SARA.
These results demonstrate that SARA is a specific partner of receptor-
regulated Smads in the TGF(3/activin signaling pathway and further suggest
that
TGF(3 signaling induces dissociation of SARA/Smad complexes.
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
44
The present invention is not limited to the features of the embodiments
described herein, but includes all variations and modifications within the
scope
of the claims.
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
REFERENCES
Abdollah, S., Macias-Silva, M., Tsukazaki, T., Hayashi, H., Attisano, L.,
and Wrana, J.L. (1997). Tf~RI phosphorylation of Smad2 on Ser 465 and 467 is
5 required for Smad2/Smad4 complex formation and signaling. J. Biol. Chem.
272,
2 76 78-2 7685.
Attisano, L. and Wrana, J.L. (1998). Mads and Smads in TGFf~ signaling.
Curr. Op. Cell Biol. J0, 188-194.
Attisano, L., Wrana, J.L., Cheifetz, S., and Massague, J. (1992). Novel
activin receptors: Distinct genes and alternative mRNA splicing generate a
repertoire of serine/threonine kinase receptors. Cell 68, 97-108.
15 Bean, A.J., Siefert, R., Chen, Y.A., Sacks, R., and Scheller, R.H. (1997).
Hrs-2 is an ATPase implicated in calcium-regulated secretion. Nature 385, 826-
829.
Burd et al., (1998), Mol. Cell., 2, 157-162.
Chen, H.I. and Sudol, M. (1995). The WW domain of Yes-associated
protein binds a proline-rich ligand that differs from the consensus
established for
Src homology 3-binding modules. Proc Natl Acad Sci USA 82, 7819-7823.
Chen, X., Rubock, M.J., and Whitman, M. (1996). A transcriptional
partner for MAD proteins in TGF-f~ signaling. Nature 383, 691-696.
Chen, X., Weisberg, E., Fridmacher, V., Watanabe, M., Naco, G., and
Whitman, M. (1997a). Smad4 and FAST-1 in the assembly of activin-responsive
factor. Nature 389, 85-89.
Chen, Y., Bhushan, A., and Vale, W. (1997b). Smad8 mediates the
signaling of the receptor serine kinase. Proc. Natl. Acad. Sci. USA 94, 12938-
12943.
Chen, Y., Lebrun, J.-J., and Vale, W. (1996). Regulation of transforming
growth factor f~- and activin-induced transcription by mammalian Mad proteins.
Proc. Natl. Acad. Sci. USA 93, 12992-12997.
40 Dennler, S., Itoh, S., Vivien, D., ten Dijke, P., Huet, S., and Gauthier,
J.-
M. (1998). Direct binding of Smad3 and Smad4 to critical TGF(3-inducible
elements in the promoter of human plasminogen activator inhibitor-type 1 gene.
EMBO J. 17, 3091-3100.
45 Dyson, S. and Gurdon, J.B. (1998). The Interpretation of Position in a
Morphogen Gradient as Revealed by Occupancy of Activin Receptors. Cell 93,
557-568.
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
46
Faux, M. and Scott, J.D. (1996). Molecular glue: kinase anchoring and
scaffold proteins. Cell 85, 9-12.
Gaullier et al., (1998), Nature, 394, 433-434.
Heldin, C.-H., Miyazono, K., and ten Dijke, P. (1997). TGF-(3 signaling
from cell membrane to nucleus through SMAD proteins. Nature. 390, 465-471.
Henis, Y.I., Moustakas, A., Lin, H.Y., and Lodish, H.F. (1994). The type II
and III transforming growth factor-f3 receptors form homo-oligomers. J. Cell
Biol.
126, 139-154.
Hoodless, P.A., Haerry, T., Abdollah, S., Stapleton, M., O'Connor, M.B.,
Attisano, L., and Wrana, J.L. (1996). MADR1, a MAD-related protein that
functions in BMP2 signaling pathways. Cell 85, 489-500.
Janknecht et al., (1998), Genes & Development, 12, 2114-2119.
Kim, J., Johnson, K., Chen, H.J., Carroll, S., and Laughon, A. (1997).
Drosophila Mad binds to DNA and directly mediates activation of vestigial by
decapentaplegic. Nature 388, 304-308.
Komada, M. and Kitamura, N. (1995). Growth factor-induced tyrosine
phosphorylation of Hrs, a novel 115-kilodalton protein with a structurally
conserved putative zinc finger domain. Mol Cell Biol 15, 6213-6221.
Kretzschmar, M., Liu, F., Hata, A., Doody, J., and Massague, J. (1997).
The TGF-f~ family mediator Smad1 is phosphorylated directly and activated
functionally by the BMP receptor kinase. Genes Dev. 11, 984-995.
Kretzschmar, M. and Massague, ). (1998). SMADs: mediators and
regulators of TGF-~3 signaling. Current Opinion in Genetics & Development 8,
103-111.
Labbe, E., Silvestri, C., Hoodless, P.A., Wrana, J.L., and Attisano, L.
(1998). Smad2 and Smad3 positively and negatively regulate TGF(3-dependent
transcription through the forkhead DNA binding protein, FAST2. Molecular Cell
in press.
Lagna, G., Hata, A., Hemmati-Brivanlou, A., and Massague, J. (1996).
Partnership between DPC4 and SMAD proteins in TGF-fi signaling pathways.
Nature 383, 832-836.
Liu, F., Pouponnot, C., and Massague, J. (1997a). Dual role of the
Smad4/DPC4 tumor suppressor in TGFj3-inducible transcriptional complexes.
Genes & Development 1 J, 3157-3167.
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
47
Liu, X., Sun, Y., Constantinescu, S.N., Karam, E., Weinberg, R.A., and
Lodish, H.F. (1997b). Transforming growth factor ~3-induced phosphorylation of
Smad3 is required for growth inhibition and transcriptional induction in
epithelial cells. Proc. Natl. Acad. Sci. USA 94, 10669-10764.
Macias-Silva, M., Abdollah, S., Hoodless, P.A., Pirone, R., Attisano, L.,
and Wrana, J.L. (1996). MADR2 is a substrate of the TGF(~ receptor and its
phosphorylation is required for nuclear accumulation and signaling. Cell 87,
12 i 5-1224.
Mu, F.T., Callaghan, J.M., Steele-Mortimer, O., Stenmark, H., Parton,
R.G., Campbell, P.L., McCluskey, J., Yeo, J.P., Tock, E.P., and Toh, B.H.
(1995).
EEA1, an early endosome-associated protein. EEA1 is a conserved alpha-helical
15 peripheral membrane protein flanked by cysteine "fingers" and contains a
calmodulin-binding IQ motif. J Biol Chem 270, 13503-13511.
Nakao, A., Imamura, T., Souchelnytskyi, S., Kawabata, M., Ishisaki, A.,
Oeda, E., Tamaki, K., Hanai, J.-i., Heldin, C.-H., Miyazono, K., and ten
Dijke, P.
20 (1997a). TGF-f~ receptor-mediated signaling through Smad2, Smad3 and Smad4.
EMBO J. 16, 5353-5362.
Nakao, A., Roijer, E., Imamura, T., Souchelnytskyi, S., Stenman, G.,
Heldin, C.-H., and ten Dijke, P. (1997b). Identification of Smad2, a human Mad
25 related protein in the transforming growth factor (3 signaling pathway. J.
Biol.
Chem. 272, 2896-2900.
Nakayama, T., Snyder, M.A., Grewal, S.S., Tsuneizumi, K., Tabata, T.,
and Christian, J.L. (1998). Xenopus SmadB acts downstream of BMP-4 to
30 modulate its activity during vertebrate embryonic patterning. Development
125,
857-867.
Nishimura, R., Kato, Y., Chen, D., Harris, S.E., Mundy, G.R., and Yoneda,
T. (1998). SmadS and DPC4 are key molecules in mediating BMP-2-induced
35 osteoblastic differentiation of the pluripotent mesenchymal precursor cell
line
C2C12. j. Biol. Chem. 273, 1872-1879.
Pasteris, N.G., Cadle, A., Logie, L.J., Porteous, M.E., Schwartz, C.E.,
Stevenson, R.E., Glover, T.W., Wilroy, R.S., and Gorski, J.L. (1994).
Isolation
40 and characterization of the faciogenital dysplasia (Aarskog-Scott
syndromeygene:
a putative Rho/Rac guanine nucleotide exchange factor. Cell 79, 669-678.
Patki et al., (1998), Nature, 394, 433-4.
45 Patterson, G., Koweek, A., Wong, A., Liu, Y., and Ruvkun, G. (1997). The
DAF-3 Smad protein antagonizes TGF-f3-related receptor signaling in the C.
elegans dauer pathway. Genes and Dev.
CA 02335346 2001-O1-18
WO 00/05360 PC'T/CA99/00656
48
Pawson, T. and Scott, J.D. (1997). Signaling through scaffold, anchoring
and adaptor proteins. Science 278, 2075-2080.
5 Piper, R.C., Cooper, A.A., Yang, H., and Stevens, T.H. (1995). VPS27
controls vacuolar and endocytic traffic through a prevacuolar compartment in
Saccharomyces cerevisae. J Cell Biof 131, 603-617.
Savage, C., Das, P., Finelli, A., Townsend, S., Sun, C., Baird, S., and
10 Padgett, R. (1996). The C. elegans sma-2, sma-3 and sma-4 genes define a
novel
conserved family of TGF-f~ pathway components. Proc. Natl. Acad. Sci. USA 93,
790-794.
Sekelsky, J.J., Newfeld, S.J., Raftery, L.A., Chartoff, E.H., and Gelbart,
15 W.M. (1995). Genetic characterization and cloning of Mothers against dpp, a
gene required for decapentaplegic function in Drosophila melanogaster.
Genetics 139, 1347-1358.
25
Simonsen et al., (1998), Nature, 394, 494-495.
Souchelnytskyi, S., Tamaki, K., Engstrom, U., Wernstedt, C., ten Dijke, P.,
and Heldin, C.-H. (1997). Phosphorylation of Ser465 and Ser467 in the C
Terminus of Smad2 Mediates Interaction with Smad4 and Is Required for
Transforming Growth Factor-~i Signaling. J. Biol. Chem. 272, 28107-28115.
Topper, J.N., Cai, )., Qiu, Y., Anderson, K.R., Xu, Y.-Y., Deeds, J.D.,
Feeley, R., Gimeno, C.J., Woolf, E.A., Tayber, O., Mays, G.G., Sampson, B.A.,
Schoen, F.J., Gimbrone, M.A.J., and Falb, D. (1997). Vascular MADs: two novel
MAD-related genes selectively inducibfe by flow in human vascular
endothelium. Proc. Natl. Acad. Sci. USA 94, 9314-9319.
Weisman, L.S. and Wickner, W. (1992). Molecular characterization of
VAC1, a gene required for vacuole inheritance and vacuole protein sorting. J
Biol Chem 267, 618-623.
Wiedemann et al., (1998), Nature, 394, 426-427.
Wieser, R., Wrana, J.L., and Massague, J. (1995). GS domain mutations
that constitutively activate Tf~R-I, the downstream signaling component in the
TGF-(3 receptor complex. EMBO J. 14, 2199-2208.
Wrana, J.L., Attisano, L., Carcamo, J., Zentella, A., Doody, J., Laiho, M.,
Wang, X.-F., and Massague, J. (1992). TGF-f~ signals through a heteromeric
protein kinase receptor complex. Cell 71, 1003-1014.
Wrana, J.L., Attisano, L., Wieser, R., Ventura, F., and Massague, J. (1994).
Mechanism of activation of the TGF-f~ receptor. Nature 370, 341-347.
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
49
Yamamoto, A., DeWald, D.B., Boronenkov, I.V., Anderson, R.A., Emr,
S.D., and Koshland, D. (i 995). Novel PI(4)P S-kinase homologue, Fab1 p,
essential for normal vacuole function and morphology in yeast. Mol Biol Cell
6,
S 525-539.
Yingling, J.M., Datto, M.B., Wong, C., Frederick, J.P., Liberati, N.T., and
Wang, X.-F. (1997). Tumour Suppressor Smad4 is a Transforming Growth Factor
~i-Inducible DNA Binding Protein. Mol. Cell. Biol. 17, 7019-7028.
Zawel, L., Dai, J.L., Buckhaults, P., Zhou, S., Kinzler, K.W., Vogelstein,
B., and Kern, S.E. (1998). Human Smad3 and Smad4 are sequence-specific
transcription activators. Mol. Cell 1, 611-617.
15 Zhang, Y., Musci, T., and Derynck, R. (1997). The tumor suppressor
Smad4/DPC4 as a central mediator of Smad function. Curr. Biol. 7, 270-276.
CA 02335346 2001-O1-18
w0 00/05360 f~C.T; CA99/00656
SO
TABLE 1 - hSARAl - SoQUence ID NO~1
GCATACTGAATCAGCAGGACTGGCTGGTGGTGCAGCAGACATCATGAGTAAGCACCGA
GAAGTCTGTTCCTTATCACGTGTGTAAGGGGAAAAAGGIiTAAACAAGTCTCTTAAGT
GGTGTTTCCTCACCGATGGAGAATTACTTCCAAGCAGAAGCTTACAACCTGGGACAAG
GTGTTAGATGAATTTGAACAAAACGAAGATGAAACAGTTTCTTCTACTTTATTGGATA
CAAAGTGGAATAAGATTCTAGATCCCCCTTCTCACCGGCTGTCATTTAACCCTACTTT
GGCCAGTGTGAATGAATCTGCAGTTTCTAATGAGTCACAACCACAACTGAAAGTCTTC
TCCCTGGCTCATTCAGCTCCCCTGACCACAGAGGAAGAGGATCACTGTGCTAATGGAC
AGGACTGTAATCTAAATCCAGAGATTGCCACAATGTGGATTGATGAAAATGCTGTTGC
AGAAGACCAGTTAATTAAGAGAAACTATAGTTGGGATGATCAATGCAGTGCTGTTGAA
GTGGGAGAGAAGARATGTGGAAACCTGGCTTGTCTGCCAGATGAGAAGAATGTTCTTG
TTGTAGCCGTCATGCATAACTGTGATAAAAGGACATTACAAAACGATTTACAGGATTG
TAATAATTATAATAGTCAATCCCTTATGGATGCTTTTAGCTGTTCACTGGATAATGAA
AACAGACAAACTGATCAATTTAGTTTTAGTATAAATGAGTCCACTGAAAAAGATATGA
ATTCAGAGAAACAAATGGATCCATTGAATAGACCGAAAACAGAGGGGAGATCTGTTAA
CCATCTGTGTCCTACTTCATCTGATAGTCTAGCCAGTGTCTGTTCCCCTTCACAATTA
AAGGATGACGGAAGTATAGGTAGAGACCCCTCCATGTCTGCGATTACAAGTTTAACGG
TTGATTCAGTAATCTCATCCCAGGGAACAGATGGATGTCCTGCTGTTAAAAAGCAAGA
GAACTATATACCAGATGAGGACCTCACTGGCAAA,ATCAGCTCTCCTAGGACAGATCTA
GGGAGTCCAAATTCCTTTTCCCACATGAGTGAGGGGATTTTGATGAAA,A.AAGAGCCAG
CAGAGGAGAGCACCACTGAAGAATCCCTCCGGTCTGGTTTACCTTTGCTTCTCAAACC
AGACATGCCTAATGGGTCTGGAAGGAATAATGACTGTGAACGGTGTTCAGATTGCCTT
GTGCCTAATGAAGTTAGGGCTGATGAAAATGAAGGTTATGAACATGAAGAAACTCTTG
GCACTACAGAATTCCTTAATATGACAGAGCATTTCTCTGAATCTCAGGACATGACTAA
TTGGAAGTTGACTAAACTAAATGAGATGAATGATAGCCAAGTAAACGAAGAAAAGGAA
AAGTTTCTACAGATTAGTCAGCCTGAGGACACTAATGGTGATAGTGGAGGACAGTGTG
TTGGATTGGCAGATGCAGGTCTAGATTTAAAAGGAACTTGCATTAGTGAAAGTGAAGA
ATGTGATTTCTCCACTGTTATAGACACACCAGCAGCAAATTATCTATCTAATGGTTGT
GATTCCTATGGAATGCAAGACCCAGGTGTTTCTTTTGTTCCAAAGACTTTACCCTCCA
AAGAAGATTCAGTAACAGAAGAAAAAGAAATAGAGGAAAGCAAGTCAGAATGCTACTC
AAATATTTATGAACAGAGAGGAAATGAGGCCACAGAAGGGAGTGGACTACTTTTAAAC
AGCACTGGTGACCTAATGAAGAAAAATTATTTACATAATTTCTGTAGTCAAGTTCCAT
CAGTGCTTGGGCAATCTTCCCCCAAGGTAGTAGCAAGCCTGCCATCTATCAGTGTTCC
TTTTGGTGGTGCAAGACCCaAGCAACCTTCTAATCI'TAAACT'~CAAATTCCAAAGCCA
TTATCAGACCATTTACAAAATGACTTTCCTGCAAACAGTGGAAATAATACTAAAAATA
AAAATGATATTCTTGGGAAAGCAAAATTAGGGGAAAACTCAGCAACCAATGTATGCAG
TCCATCTTTGGGAAACATCTCTAATGTCGATACAAATGGGGAACATTTAGAAAGTTAT
GAGGCTGAGATCTCCACTAGACCATGCCTTGCATTAGCTCCAGATAGCCCAGATAATG
ATCTCAGAGCTGGTCAGTT'rGGAATTTCTGCCAGAAAGCCATTCACCACGCTGGGTGA
GGTGGCTCCAGTATGGGTACCGGATTCTCAGGCTCCAAATTGCATGAAATGTGAAGCC
AGGTTTACATTCACCAAAAGGAGGCATCACTGCAGAGCATGTGGGAAGGTTTTCTGTG
CTTCCTGCTGTAGCCTGAAATGTAAACTGTTATACATGGACAGAAAGGAAGCTAGAGT
GTGTGTAATCTGCCATTCAGTGCTAATGAATGCTCAAGCCTGGGAGAACATGATGAGT
GCCTCAAGCCAGAGCCCTAACCCTAACAATCCTGCTGAATACTGTTCTACTATCCCTC
CCTTGCAGCAAGCTCAGGCCTCAGGAGCTCTGAGCTCTCCACCTCCCACTGTGATGGT
ACCTGTGGGAGTTTTAAAGCACCCTGGAGCAGAAGTGGCTCAGCCCAGAGAGCAGAGG
CGAGTTTGGTTTGCTGATG.~GATCTTGCCCAATGGAGhAGTTGCTGATGCAGCCAAAT
TAACAATGAATGGAACTTCCTCTGCAGGAACCCTGGCTGTGTCACrICG.".sCCCAGTCAA
GCCAGTAACTACCAGTCC~'CTr.CCAGCAGAGACGGATATTTGTCTATTCTCTGGGAGT
ATAACTCAGGTTGGAAGTCCTGTTGGAAGTGCAATGAATCTTATTCC
TGAAGATGGCC
TTCCTCCCATTCTCATCTCCACTGGTGTAA.AAGGAGACTATGCTG:GGA~,GAGAAACC
ATCACAGATTTCAG T iiATG C:-.GC.~GTTGGAGGATGG i G:~ C. C C : ;~AC C
C.'~CTTGTATTT
GTTTTAAATGCAAATT:GTTGTCAATGGTTAAAATTGT'~.TTATGTG~CAGGAAGT
GCTGGTGTTTCACAACCAAGGGAATGCATGCAGTGGG T C:~G ~ ~':'~.=";.:,T:.~TCATTCT
TCTACAGTGTTTAC C;~G.:..TG:~.AAGTG T TTG C CAAAG V,~-, y:-~ T C =""'"n~ T CAC T
TTGTG
CAGCT':'TATCGGG.=.~ "C ~'~ ~-,.,.- ..,....., ..,.... ..- . .. ,....,_, ~"
.,..,C~rZ~CT
..aV't~VVV~TG1VVIGsaV~ru~'~T.aVVt~a\.rall~ i
'='CAGTCAAAG""""'~~'""'~~~_~~-~.,ppGAACATG~'~G~,"' '- ..-._.."...,.._ ",
li-'w-JV~Vl Vi VCflT T~it'SlVIVtILt~lCiAC
SUBSTITUTE SHEET (RULE 26)
CA 02335346 2001-O1-18
WO OOI05360 PCTlCA99/00656
~1
TABLE 1 - hSARAl Continued
CTACCAGTCACTGCAAGACCTAGTACTCCCAACCCCACCTTACTTGTT.VVVIfiTV.~T
ATCCAGAAATGGGAAACTCCTTGGGCTAAAGTATTTCCTATCCGTCT~~-.TGTTGAGAC
TTGGAGCTGRATATCGACTTTATCCATGCCCACTATTCAGTGTCAGATT:CGGAAGCC
ATTGTTTGGAGAGACGGGGCATACCATCATGAATCTTCTTGCAGACTTC AG'r.~,ATTr.C
CAGTATACCTTGCCAGTAGTTCAAGGTTTGGTGGT?GATATGGAAGTTCGG:~AAACTA
GCATCAAAATTCCCAGCAACAGATACAATGAGATGATGAAAGCCATGAACAAGTCCAA
TGAGCATGTCCTGGCAGGAGGTGCCTGCTTCAATGAAAAGGCAGACTC~CATCTTGTG
TGTGTACAGAATGATGATGGAAACTATCAGACCCAGGCTATCAGTATTCACAATCAGC
CCAGAAAAGTGACTGGTGCCAGTTTCTTTGTGTTCAGTGGCGCTCTGAAATCC:C'r'TC
TGGATACCTTGCCAAGTCCAGTATTGTGGAAGATGGTGTTATGGTCCAGr.TTACTGCA
GAGAACATGGATTCCTTGAGGCAGGCACTGCGAGAGATGAAGGACTTCACCATCACCT
GTGGGAAGGCGGACGCGGAGGAACCCCAGGAGCACATCCACATCCAGTGG.GTGGATGA
TGACAAGAACGTTAGCAAGGGTGTCGTAAGTCCTATAGATGGGAAGTCCATGGAGACT
ATAACAAATGTGAAGATATTCCATGGATCAGAATATAAAGCAAATGGAAAAGTAATCA
GATGGACAGAGGTGTTTTTCCTAGAAAACGATGACCAGCACAATTGCCTCAGTGATCC
TGCAGATCACAGTAGATTGACTGAGCATGTTGCCAAAGCTTTTTGCC~TGCTCTCTGT
CCTCACCTGAAACTTCTGAAGGAAGATGGAATGACCARACTGGGACTACGTGTG:,CAC
TTGACTCAGATCAGGTTGGCTATCAAGCAGGGAGCAATGGCCAGCCCCT':'CCCTCGCA
GTACATGAATGATCTGGATAGCGCCTTGGTGCCGGTGATCCATGGAGGGGCCTGCCAG
CTTAGTGAGGGCCCCGTTGTCATGGAACTCATCTTTTATATTCTGGAAAACATCGTAT
AAACAGAGAAGACTTCATTTTTTTCTGTTCAGACTTGTTGCAACAGCAGTCATACCCA
AATCATTTGCACTTTAAAACTGGAAGATTAAGCTTTTGTTAACACTATTAATGGGGTG
GGGAATAGGGTGGGAGTGGGGGTTTGGGAGACGGGTGGGAAAGGGTGGTTGGGGGGAC
CGATGTTCCATAATTCTAAGTCTTCTATGCATTGTCCACCAAGAAGATCTGGGCAGCT
TCTGTTCCTGCACAACAGTTATGCTATCCTTGCAGCTAATCCCCTTCTGTTACTGTTT
AGACAAGAATTCCGCTCCTCTCTCAAGATTTACTTATGGTCATGTGCTCAGAAATGCT
CAAATGGGTACAACCATCACCAAGGGTGGGATGGGAGGGCAGAGGGGAAATAAAATAT
AAAGCAT AAAA.AAA
SUBSTITUTE SHEET (RULE 26)
CA 02335346 2001-O1-18
WO 00/05360 PCT/CA99/00656
52
TAHL$ 2 hSJIRA1 - SeQUence ID NO~2
MWIDENAVAEDQLIKRNYSWDDQCSAVEVGEKKCG1TLACLPDEKNVLVVAVMHNCDKR
TLQNDLQDCNNYNSQSLMDAFSCSLDNENRQTDQFSFSINESTEKDMNSEKQMDPLNR
PRTEGRSVNHLCPTSSDSLASVCSPSQLKDDGSIGRDPSMSAITSLTVDSVISSQGTD
GCPAVKRQENYIPDEDLTGICISSPRTDLGSPNSFSHMSEGILMKKEPAEESTTEESLR
SGLPLLLXPDMPNGSGRNNDCERCSDCLVPNEVRADENEGYEHEETLGTTEFLNMTEH
FSESQDMTNWKLTKLNEMNDSQVNEEKEKFLQISQP~DTNGDSGGQCVGLADAGLDLK
GTCISESEECDFSTVIDTPAANYLSNGCDSYGMQDPGVSFVPKTLPSKEDSVTEEKEI
EESRSECYSNIYEQRGNEATEGSGLLLNSTGDLMKIQSYLHNFCSQVPSVI~GQSSPKVV
ASLPSISVPFGGARPKQPSNLKLQIPFtPLSDHLQNDFPANSGNNTKNKNDILGKAKI,G
ENSATNVCSPSLGNISNVDTNGE$LESYEAEISTRPCLALAPDSPDNDLRAGQFGISA
RKPFTTLGEVAPVWVPDSQAPNCMKCEARFTFTKRRHHCRACGKVFCASCCSLKCKLL
Y1~RKEARVCVICHSVLMNAQAW~NI~ZSASSQSPNPNNPAEYCSTIPPLQQAQASGAL
SSPPPTVMVPVGVLKIiPGAEVAQPREQRRVWFADGILPNGEVADAAKLTMNGTSSAGT
LAVSHDPVKPVTTSPLPAETDICLFSGSITQVGSPVGSAMNLIPEDGLPPILISTGVK
GDYAVEEKPSQISVMQQLEDGGPDPLVFVLNANLLSMVKIV'NYVN'RKCWCFTTKGMHA
VGQSEIVILLQCLPDEKCLPRnIFNHFVQLYRDALAGNWSNLGHSFFSQSFLGSKEH
GGFLYVTSTYQSLQDLVLPTPPYLFGILIQKWETPWAKVFPIRLMLRLGAEYRLYPCP
LFSVRFRKPLFGETGHTIMNLLADFRNYQYTLPVVQGLVVDMEVRKTSIKIPSNRYNE
MMg:AMNKSNEHVLAGGe'~CFNERADSFILVCVQNDDGNYQTQAI S IHNQPRKVTGASFFV
FSGALKSSSGYLAKSSIVEDGVMVQITAENMDSLRQALREMKDFTITCGKADAEEPQE
HIFiIQWVDDDKNVSKGWSPIDGKSMETITNVKIFHGSEYKANGRVIRWTEVFFLEND
DQHNCLSDPADHSRLTEHVAKAFCLALCTQLKLLKGDGMTF~,LGLRVTLDSDQVGYQAG
SNGQHLPSQYMNDFDSDLVKMIHGGACQLSEGPVVMELIFYILENIV
SUBSTITUTE SHEET (RULE 26)
CA 02335346 2001-O1-18
WO 00/05360 PCTlCA99/00656
53
TJ18LE 3 human SARA2 Seauence ID NO~3
ACTCCCGGCCGGGGTAGCTC':"-'CACTCC'r'CAGCGCGACGTCGTG T CGAG T T C C Ca,AA.A
AGCTCCGCAGGGGCTGTAGGGAGGTGATCTCATCCATTRACAGCTGT.~TGTTGCCAGT
TCCCAAATCTTTATCTATCTCAGACTTCTCTCCTGCATTCCAGATTC'-"W.T:.T TCAGC
TGCCTTTTGGATATCTCTCCCAGGATGTTCTCAAGGCATACAAGAATTAAATTCTGAA
TAAGTCTGCAGGTAGGATGGACAGTTATTTTAAAGCAGCTGTCAGTG~.C:TGGr.CAAA
CTCC:TGATGATTTTGAACAGAACCCAGATGAACAAGATTATCTC~uCAGATGTACAAA
ATGCATATGATTCTAACCACTGCTCAGTTTCTTCAGAGTTGGCTTCCTCACAGCGAAC
TTCATTGCTCCCAAAAGACCAAGAGTGCGTTAATAGTTGTGCCTCATCAGAAACAAGC
TATGGAACAAATGAGAGTTCCCTGAATGAAAAAACACTCAAGGGACTTr.CT'.'CTATAC
AAAATGAAAAAAATGTAACAGGACTTGATCTTCTTTCTTCTGTGGATGGTGGTACTTC
AGATGAAATCCAGCCGTTATATATGGGACGATGTAGTAAACCTATCTGTGATCTGATA
AGTGACATGGGTAACTTAGTTCATGCAACCAATAGTGAAGAAGATATT~,AAAAATTAT
TGCCAGATGATTTTAAGTCTAATGCAGATTCCTTGATTGGATTGGATTTATCTTCAGT
GTCAGATACTCCCTGTGTTTCTTCAACAGACCATGATAGTGATACTGTCAGAGAACAA
CAGAATGATATCAGTTCTGAATTACAAAATAGAGAAATCGGAGGAATCAAAGAATTGG
GTATAAAAGTAGATACAACACTTTCAGATTCCTATAATTACAGTGGAAC:-~Ga.AAATTT
AAAAGATAAAAAGATCTTTAATCAGTTAGAATCAATTGTTGATTTTAACATGTCATCT
GCTTTGACTCGACAAAGTTCCAAAATGTTTCATGCCAAAGACAAGCTACAACACAAGA
GCCAGCCATGTGGATTACTAAAAGATGTTGGCTTAGTAAAAGAGGAAGTAGATGTGGC
AGTCATAACTGCCGCAGAATGTTTAAAAGAAGAGGGCAAGACAAGTGCTTTGACCTGC
AGCCTTCCGAAAAATGAAGATTTATGCTTAAATGATTCAAATTCAAGAGATG~.A.A,ATT
TCAAATTACCTGACTTTTCCTTTCAGGAAGATAAGACTGTTATAAAACAATCTGCACA
AGAAGACTCAAAAAGTTTAGACCTTAAGGATAATGATGTAATCCAAGATTCCTCTTCA
GCTTTACATGTTTCCAGTAAAGATGTGCCGTCCTCATTGTCCTGTCTTCCTGCGTCTG
GGTCTATGTGTGGATCATTAATTGAAAGTAAAGCACGGGGTGATTTTTTACCTCAGCA
TGRACATAAAGATAATATACAAGATGCAGTGACTATACATGAAGAAATACAGAACAGT
GTTGTTCTAGGTGGGGAACCATTCAAAGAGAATGATCTTTTGAAACAGGAAAA.ATGTA
AAAGCATACTCCTTCAGTCATTAATTGAAGGGATGGAAGACAGAAAGATAG~.TCCTGA
CCAGACAGTAATCAGAGCTGAGTCTTTGGATGGTGGTGACACCAGTTCTACAGTTGTA
GAATCTCAAGAGGGGCTTTCTGGCACTCATGTCCCAGAGTCTTCTGATTGTTGTGAAG
GTTTTATTAATACTTTTTCAAGCAATGATATGGATGGGCAAGACT ~'AGasT T.=~;.'~"~'TAA
TATTGATGAAGGCGCAAAAAGTGGCCChCTAATTAGTGATGCTGAAC:'~G~TGCCTTT
CTGACAGAACAGTATCTTCAGACCnCT?.ACATAAAGTCTTTTGAAGAA.AATGTAAATG
ACTCTAAATCGCAAATGAATCAGATAGATATGAAAGGCTTAGATGATG:~~CATCAA
TAATATATATTTCAATGCAGAAGCAGGAGCTATTGGGGAAAGTCATGGTATTa.ATATA
ATTTGTGAAACAGTTGATAAACAAAATACAATAGAAAATGGCCTTTCTT"'AGGAGAAA
AAAGCACTATTCCAGTTCAACAAGGGTTACCTACCAGTAAGTCTGAGaTTA.:AAATCA
ATTATCAGTCTCTGATATTAACAGTCAATCTGTTGGAGGGGCCAGACCTAAGC.yATTG
TTTAGCCTTCCATCAAGAACAAGGAGTTCAAAGGACCTGAATAAGCCAGe'~.T.;TTCCAG
ATACAATAGRAAGTGAACCCAGCACAGCAGATACCGTTGTTCCAATCACTT~TGCTAT
AGATTCTACAGCTGATCCACAGGTTaGCT'~'CAACTCTAATTACATTGATAT~-.GAAAGT
AATTCTGAAGGTGGATCTAGTTTCGThACTGCAAATGAAGATTCTGTaCCTGAA.A.ACA
CTTGCAAAGAAGGCTTGGTTTTGGGC C:~GAAACAGCCTACTTGGG T T C C C',G:.~ T TCAGA
AGCTCCAAACTGTATGAACTGCCAAGTC«AATTTACTTTTACCAAACGGCG~CaCCAT
TGCCGAGCATGTGGGAAAGTAT T"'TG T GGTGTCTGTTGTAATAGG~.AG T G':'~ACTGC
AATATCTAGAAAAGGAAGCAAGAGT:~TGTGTAGTCTGCTATGAAACTATTyG:nAAGC
TCAGGCAT?TGAAAGGATGATG:..G : C ~~,CTGGTTCTAATCTTAAGT C':'~T CnT TC T
GATG~T G TACTACTGTCCAGCC ~' C C':" _AGGAGAACCAAACATC CnG T n T:~;. C'_ TCAC
CAGG"yACTT T GCCAGTCTCAGCr".CT ~'W~.AACCAGGTGTTGAAG~hC'~':~ T u'_"_'.~. CAA
AGi.~:yC.=.G:~Gr'1GAGTATGG T T .'~CW,.=.T~~s TATATTGCCCAATG" i.~:~i.."'-
='.;Cr.GAT
ACAA "C.~AAATTATCATCTGGAhG T ~,~,G~,TG T T CTGAAGAC':"_'"'hG T C C'" C "'
C~'CAC
CTGA T G T G C C =ATGACAG 1 wW/'f Vfl V 1 V V llT C!'~ l TC v. ~TT
CTII'T~'iL.tf V Z V V
GC Cra~u~CnATGe~GACAGGisv;si~ T:z i ~":W.i~iwe~ae~~ATGe"~aGATAATT 'raGi-~G'~'
_ = =' ~~ T TAT
...._.~..,._ ~.GGTGGAi.~.'" .. ; _
v.AVV 1 w.~l ...~_ m1 I~ ~~~ n7~ ~_ .~.~.-.~ .. CTw.
1 ~3T.~~ly~ VMl..t1't'fVV~TV/9V V V V~~..~ ~ ~~
..TGVTTriaTTTACACTAG ~""'~"" ~~~~'~ T~w~~_~~---_ ,.,
lllGCilVtll'Twww '~V7~.l1V~C1VMV1u1\.v:.~.~~aGiL.
SUBSTITUTE SHEET (RULE 26)
CA 02335346 2001-O1-18
WO OOI05360 PCTlCA99100656
54
TAEZE 3 human SARA2 - Continued
TACTGG T G T ~':"='~,GTTAACAGCAATTTACC~.=.T T G ~ T:,GTATTTCAGAT T A ~AGG T':
a,
CTGTGTGATATTAACAAGTATGTCTGCAATAAGnTT~.GTCTTCTACCTAATGATGr.GG
ACAGT T T GC C C C CACTTCTGGTTGCATCTGGAGA~,AAGGGATCAGTGCC T G T AGT:~G.=.
AGAACATCCATCTCATGAGCAGATCATTTTGCT'~C-=GAAGGTGAAGGCT::'C:.TCC~'
GTTACATTTGTCCTAAATGCTAATCTACTCGTGAATGTCAAATTCATATTTTATTCC:
CAGACAAATATTGGTACTTTTCAACCAATGGATTGCATGGCTTGGGACAGGCAGAAAT
TATTATTCTATTGTTATGTTTGCCAAATGAAGATACTATTCCTRAGGACATCTTCAGA
CTATTTATCACCATA?ATAAGGATGCTCTAAAAGG.=~AAATACATAGARAACTT~vGAC.~
ATATTACCy~ ACTGAGAGTTTTCTCAGTAGCAAGGATCACGGAGGATTCCTGTTTAT
TACACCTACTTTTCAGAAACTTGATGATCTCTCATTACCAAGTAATCCTTTTCTTTGT
GGAATTCTTATCCAGAAGCTTGAGATTCCCTGGGCAAAGGTTTTTCCTATGCGTTTAA
TGTTGAGATTGGGTGCAGAATATAAAGCATATCCTGCTCCTCTAACAAGCATCAGAGG
CCGAAAACCTCTTTTTGGAGRAATAGGACACACTATTATGAACTTACTTGTTGACCTT
CGAAATTACC.~GTATACCTTGCATAATATAGATCAACTGTTGATTCATATG~v.~.AATGG
GAAAAAGCTGCATA.AAAATACCACGGAAAAAGTACAGTGATGTAATGAAAGTACTAAA
TTCTTCCAATGAGCATGTCATTAGCATTGGAGCAAGTTTCAGTACAGAAGCAGATTCT
CATCTAGTCTGTATACAGAATGATGGAATTTATGAAACACAGGCCAACAGTGCCACTG
GCCATCCTAGAAAAGTGACAGGTGCAAGTTTTGTGGTATTCAATGGAGCTCTAAAAAC
ATCTTCAGGATTTCTTGCTAAGTCCAGCATAGTTG~,AGATGGCTTAATGGTACAAATA
ACTCCAGAGACCATGAATGGCTTGCGGCTAGCTT:ACGAGAACAGAAAGACTTTA.AA.A
TTACATGTGGG:.~AAGTTGATGCAGTAGACCTGAGAGAATACGTGGATATCTGCTGGGT
AGATGCTGAAGAAAAAGGAAACAAAGGAGTTATCAGTTCAGTGGATGGAATATCATTA
CAAGGATTTCCAAGTGAAAAAATAAAACTGGAAGCAGATTTTGAAACCGATGAGAAGA
TTGTAAAATGTACCGAGGTGTTCTACTTTCTAAAGGACCAGGATTTATCTATTTTATC
AACTTCTTATCAGTTTGCAAAAGAAATAGCCATGGCTTGTAGTGCTGCGCTGTGCCCT
CACCTGAAAACTCTAAAAAGTAATGGGATGAATAAA.ATTGGACTCAGAGTTTCCATTG
ACACTGATATGGTTGAATTTCAGGCAGGATCTGAAGGCCAACTTCTGCCTCAGCATTA
TCTAAATGr'1TCTTGATAGTGCTCTGATACCTGTGhTCCATGGTGGGACCTCCAACTCT
AGTTTACCATTAGAAATAGAATTAGTGTTTTTCATTATAGAACATCTTTTTTAGTGAA
AGAATGTGCCnTATTACATATTGCAACCTAATT';'GTTAAAACTAACTCCAGCACTAAA
GCTGAAATGCCACnA.ACACTAAAAGTATAAATATGTCr'GATTTTTGAAACACATAAGC
TTTGCTC':"_'TAGGCAGGAATGATCTTTTCAAATCAT T:-.GCACAATATTTAAATATCT.-.
AAAATTTAAGAGhTCCATACTTTCTGTAGCT'=' T ACAi..TTAATTTAAGTACTAAAA.AG.=.
CAAGGATTTCT'"TTAAGAAATTTATAGCATTTACTGTGTTATTTAA.ATGCTAAGCCAA
AGTATCTGCACTTAGGTATACCTCTTTATGCCAATt.ATGATTTTAATGAAGGCTCTTT
TCAGATGTAACCTTATGAAGGAAATATCTGCTTTGTGTATATGCCAGTTAGAATACTG
GTTTCTAAAGTCTGTCAAATTGTATTTCAGTGGCACe'~AAAACCAGTTTTGAGGTCTTA
GACTTATAATT C'T_'TTGAATAAAACTGATAACTTAT ~"~ GTATAATTGGAG T G GAGAC C'.'
ACCTCCATAAT'~'AGATAAACTCTTTTTGGATTATAATCAGAATTTTGCCTTTTTTCT:'
CTCAAATTATTACATATGTATGTATTATATATCCACATATATAGTTTTCCCTGATTAA
ATGGATATTAAAATAATTGCGGGTGCTTCAGGACT~'~'TTGCTTCTATATTTAAGTAT:y
TTGTTTT':'ATAGCAAGAACATATTCTGAATGT T :':'ATAA.ATCTT TAATAATTTATATG
TAGGTAATe"~T'="_~ ~'TGTATCACAATGCATTAT T :."~'~'~'~'~CCTCCTTTCCT? ~~. CAAAC T
a,
TACCACTG T A T TTACCACTTCTAAGAGTGAC T a:~C G~=.CGGGCCAGATGACCC':'TGAAG
TAGTCA T T AT G T T.G CAATAAATGAAG C CTGAAACAG G T : '-":'T'I'TACTT C CF.C T T
T AAT
CCTTAGAi~rIT =":' ~ :'~'GGCAACTTCG CATAT'~ -"" ~~., y
"';,ew.~C'rlCT.iGTGTATAAG T ATi~A
ATTTAAATi~~-,'r.C .:...ATTACTTTTGCATAT T ~""A~ T "' C"' T TATATGGTAG T T AT
~'""~'"
TATAACAGG.-, T A T T AACF~TAAGTTAAATC C =~. T G T:~ T ~":'GAAAT T GTTACAG: ~G
C T =' ~'
CCTCT:'TAC"'"'W":,ACAGCAAAA.AAGTGGGGGGCA:nT T G TAGTCCTGTCAT':'TAAG T
TATG T AAA, T ~' =':,ATCATTATTTTGATG C':":"~ ~C:-, T TC T CATG T GThATATA T G
TTTTTG T:~T CCAC T CATATATT':' C:~AGa.A~",~,T T ~=.TGT T ~ T:-.G C C C'_'
GTTTTAAG~, T,=, T ~ATGAAG CAT C'. C~.,a,C':' ~' G;~,G~., T C:~AG T CAAAG T
.'.=, T ~,r.'
TCAGG:-. T C "'.. _ ,...,......., w7~ ...., ,.,. .. _ .....,. _ .. _ _ ...
...,.. ,..,...............
VL'1VV1 V.w.~GCIlIVVAVP'lVl'1'_ VtIVM_TT~ty~lL.l1V11 _ VVVL.t'\~ 7.
T~T~~_ n w1 _ I1 111IW 1111 w I- w TTT~
AGl1 ~ VVtIW VM1 lf~ffl l.~ VTAGitIWnVy:~MVa~a'M1 ~ VVtIVVtIVMitIT~
AGC~.~..1._..,..__ _ .._w ..,...1_ .. ." T,.T T.. _ _ _ ....... ......w n _ _
,..._ _ _
~. w ~VVMVta'1V'.itlvl C'1VMCMiV~~'1C1C~V1V~~.VVt~VtfTtl~T,-Ml~yy~
SUB'TITUTE SHEET (RULE 26)
CA 02335346 2001-O1-18
w0 00/05360 PCTlCA99/00656
TABLE 3 human SARA2 Continued
ATGGCAACCTGTTATAATTTGTGARAT:TATTGAAATGGTGTAAGATGAAA1CAATTG
CATATCAAACCCAATTTATGTTTTCTAAATATAGTGTATGTATTCTGCCATGSAAGTA
ATTGAACAGTCTTAAAATAACCAAATGGTAGAGGGCTGTTCCATGATGGGACAGCTTT
GGATTTGTTTTCATAAAATCTCTACATTCAATAAAAATTGGAATTATGTGCCTGAAGT
TTGGAGGCACATTTTGAAGT
SUBSTtTUTE SHEET (RULE 26)
CA 02335346 2001-O1-18
WO 00/OS360 PC.'tlCA99/00656
~6
TABLB 4 human SARA2 - Seauence ID N0s4
I~SYFRAAVSDLDKLLDDFEQNPDEQDYLQDVQNAYDSNHCSVSSELASSQRTCLLPK
DQECVNSCASSETSYGTNESSLNEKTLKGLTSIQNEKNVTGLDLLSSVDGG~S~~IQP
LYMGRCSKPICDLISDMGNLVHATNSEEDIKKLLPDDFKSNADSLIGLDLSSVSDTPC
VSSTDHDSDTVREQQNDTSSELQNREIGGIitELGIKVDTTLSDSYNYSGTENLKDKKI
FNQLESIVDFNMSSALTRQSSKMFHAKDKLQHKSQPCGLLKDVGLVKEEVDVAVITAA
ECLKEEGKTSALTCSLPKNEDLCLNDSNSRDENFKLPDFSFQEDKTVIKQSAQEDSKS
LDLKDNDVIQDSSSALHVSSFmVPSSLSCLPASGSMCGSLIESKARGDFLPQHEHKDN
IQDAVTIHEEIQNSVVLGGEPFKENDLLKQEKCKSILLQSLIEGMEDRKIDPDQTVIR
AESLDGGDTSSTVVESQEGLSGTBVPESSDCCEGFINTFSSNDMDGQDLDYFNIDEGA
KSGPLISDAELDAFLTEQYLQTTNIKSFEENVNDSKSQMNQIDMKGLDDGNIh'NIYFN
AEAGAZGESHGINIICETVDRQNTIENGLSLGEKSTIPVQQGLPTSKSEiTNQLSVSD
INSQSVGGARPKQLFSLPSRTRSSRDLNKPDVPDTIESEPSTADTVVPITCAIDSTAD
PQVSFNSNYIDIESNSEGGSSFVTANEDSVPENTCKEGLVLGQKQFTWVPDSEAPNCM
NCQVKFTFTRRRHFiCRACGKVFCGVCCNRKCKLQYLEKEARVCWCYETISKAQAFER
MMSPTGSNLKSNHSDECTTVQPPQENQTSSIPSPATLPVSALKQPGVEGLCSKBQKRV
WFADGILPNGEVADTTRLSSGSRRCSEDFSPLSPDVPMTVNTVDHSHSTTVEKPNNET
GDITRNEIIQSPISQVPSVEKLSMNTGNEGLC'~TSGSFTLDDDVFAETEEPSjPT,~'uLV
NSNLPIASISDY'RLLCDINKYVCNKISLLPNDEDSLPPLLVASGEKGSVPVVE=PSH
EQIILLLEGEGFHPVTFVINANLLVNVKFT__FYSSDKYWYFSTNGLHGLGQAEI=ILLL
CLPNEDTIPKDIFRLFITIYKDALKGKYIENLDNITFTESFLSSKDHGGFLFITPTFQ
KLDDLSLPSNPFLCGILIQKLEIPWAKVFPMRLMLRLGAEYKAYPAPLTS_TRGRKPLF
GEIGHTIMNLLVDLRNYQYTLHNIDQLLIHMEMGKSCIKIPRKKYSDVMKVLNSSNEH
VISIGASFSTEADSHLVCIQNDGIYETQANSATGHPRKVTGASFWFNGALKTSSGFL
AKSSIVEDGLMVQITPETMNGLRLALREQKDFKITCGKVDAVDLREYVDICWVDAEEK
GNKGVISSVDGISLQGFPSEKIKLEADFETDEKIVKCTEVFYFLKDQDLSILSTSYQF
AKEIAMACSAALCPHLKTLKSNGMNKIGLRVSIDTDMVEFQAGSEGQLLPQHYLNDLD
SALIPVIHGGTSNSSLPLEIELVFFIIEHLF
SUBSTITUTE SHEET (RULE 26)
CA 02335346 2001-O1-18
WO 00/05360 PCTlCA99/00656
57
TAHL$ 5 aSxRAl - Seauence T_D NO~5
CTGTAAGTTTGACTATGTAGGAAAGCATTTCTGTTATC=':~TGAAGTATGTTTTAGAGT
CAGACCAATAAC TAAACGGTTTTCTT TTTTTTG T T'-',:, T :"~ C C C C T CAGATGAGAC T G
CTCTCCAAAGCTATTAGATGCTAAGTGGAATCAAATC:':'AGAACCGCATTCACATAAA
GTCGCTGATAACTCCGCCCTTGACAATGTCTGTAAATCAATCATTGCTATTGAAGCTC
ATCTCAAAGTCAGGTCACCCGGCTTGTCAGCCCTTGTG:~.GGTCCACATATGTGAATGG
AGAAGTAGGTATTGTGGCACCTGAAATGCCCAAAATGGTGATAGGAGACACCATTATG
GCAGAGGATTCACTTTTTAACAACACTGGTCCCTCTGAAATTGTATGCAACCCATCTA
CTGTGGAGAGTCAAAGTTTACAAGCTTTAGATGATCAATCAGTGAATATTCACAATGA
ARAAAGTGTTCTGCTCGCTGATGGCTTTTCACCATGCAGTAGCCCCAAAAGTATTATA
AACTTTGACTGCTTGACCATGGATAACGAAATGCCTTTGCACAGTCA.AATGAGTGTTG
ATGACAATGACAAAGAAACTGTAACAATTTCAGTCCTTCCAACAATCATACAGGATAC
TAGTRACGTAAGCACAGACCCAGCTATCAATAAACCTGGCACTAAAGAACCCCATAGA
GCATTAAAGGAAACCACATCAGTTATTCTGCCTGAAATAAAGCCTTACTCCACATGTG
CTGCCCTTTCGTTTGAAAATAACAATAAGGTTCCCAGTTATCAATTAAATAATACAGA
TCTACTCAGCGTTTCACCAGTGGTTGAAGCATGTAGTGAGCAGCAGCAAAAACATACA
TCTTCCTTGCATGAAGAA,AAACTTTTTGAAGGTGTTTCTGCAACGGAGTCCTTTGCAG
CCACTGCTGCGGAAACTGTACTGGATAATGAGGCTCTCC~TAGTGCTGAATTCTTTGA
CATTGTTGTAAAGAACT:'TTCTGACTCTTGTGTGATTAATGGCGACTTGACTAAAAGT
TGTGGCCTCTCTCAAGAAAGCAATGAAAAGTTTTGTGCAAGTAAAGAGTTTGAAGGAG
GGGTAGATGCTAATGTCTTGTTGGAAA,ATGCATGTGTAGCTTATAAAGAAGCAATAGA
TTTGCCTGAAGAAAATGGAACTAATGCACCAATGTCTCTGTACAATGGGTGTGATTCC
TATGGAATGAAAAACCCAGCCGTAGCTCAARACCCAR.AGAATTTACCTTCAAAAGAAG
ATTCTGTGACAGAAG:~AAAGAAATTGAAGAAAGCAAGTCAGAATACTATACTGGTGT
TTATGAACAACAAAGAGAAGATGATGTTACAGAGAGAGGTGGACTTCTGTTAAATGCT
AAGGCTGACCAAATGAAGAACAATTTGCATAGTCTTTGTAATCAGGTTCCATCCATGC
ATGGGCAAACATCACCAAAAAAGGGCAAGATTGTGCAATCTCTCAGTGTTCCATACGG
TGGAGCACGCACTAAGCAGCCAACTCATCTCAAACTCCATATTCCAAAGCCATTGTCT
GAAATGTTGCAGAGCGATCTCATTCCTCCAAATGCTGGCTGCAGCTCTAAATACAA.AA
ATGACATGTTAAACAAATCAAATCAGGGGGATAACCTGATTTCAGAATCACTGCGTGA
GGATTCTGCAGTGCGCAGCCCTGTTACTGATGCTAATGGTGATTTCCCTGGAGAATAC
AGGGGACCTGGCAGCTTGTGCCTTGCAGTGTCTCCAGACAGCCCAGACAACGATCTGC
TTGCCGGGCAGTTTGGGGTACCCATCTCTAAGCCATC"~'r.CTACTCTAGGGGAAGTGGC
TCCAGTCTGGGTGCC:'~GATTCCCAAGCACCAAACTGCATGAAGTGCGAGGCCAGATTT
ACATTTACCAAAAGGAGGCATCACTGCCGAGCTTGTGGAAAGGTGTTCTGTGCTGCTT
GTTGCAGTCTAAAATG"..AA.ACTACAGTACATGGATAAAAAGGAGGCTCGTGTGTGTGT
TATTTGTCATTCTGTGCTTATGAATGCTCAAGCATGGGAGAACATGTTAAGTGCATCG
GTCCAAAGCCCAAATCCAAATAATCCTGCTGAATACTGCTCAACTATCCCTCCGATGC
AGCAGGCACAAGCTTCAGGAGCACTGAGTTCCCCACCTCCCACTGTCATGGTGCCAGT
GGGTGTGTTAAAACATCCAGGAACTGAAGGGTCACAGTCAAAGGAACAGCGCCGTGTT
TGGTTTGCTGATGGAATATTACCCAACGGAGAGACTGCTG.=.CTCAGATAATGCAAACG
TAACTACAGTGGCTGGGACACTTACTGTGTCACATACCAACAATTCCACATCTTCAGA
GTCTGAGAACACCTCTGGATTCTGTGGAAGTATAACTCAGGTTGGCAGTGCAATGAAC
CTTATTCCAGAAGATGGGCTTCCTCCTATACTAATCTCTa.CTGGAGTAAAAGGAGATT
ACGCAGTTGAGGAACGCCC:'TCCCAGATGTCTGTGATGCAGCAACTAGAGGAAGGAGG
ACCAGATCCTT T GG T'_"=":' T G T TCTAAATGCAAATC':'""~"'GGC Ca T GG T TAAGATCGTG
AACTATGTTAACAGGAAATGCTGGTGCTTTACTAC"~AGGGAATGCATGCAGTGGGCC
AGGCTGAGATCG THAT CC T G TTGCAGTGCCTGCC'='u AT G:..,GAAG T GCCTGCCGAGGGA
CCTGTTTAGC CA T'="""';~ T TGAGC T GTATCAGGfsGGCAA T :',~ CAG;~ T AATG TAGTGGGG
AACCTGGGGC~-. T T C C""" C ~'~ CAGCCAGAGTTTCC'_' GuG T AG T AAGGAT CATGGTGGAT
TTCTTTATGTTGCACCAACCTACCAGTCCCTCCAGGAC:.:',J"T'_CTTCCTGCAGAGCC
GTACTTGTT i V Vt~.A 1 ~ _ _ _ ~..TT~AAAAGTGGG.:.aG:~ C'.' C CA T GGG C ,.
""' " ~AAAGTGTTCCCC
ATT C GGC T T AT G C T ~;. G T T':'AGGTG CAGAATACAGA T '=' ~ .."-.C C C:~T G T
C C.=.CT CTT CA
GTGTTCGATACAG:,:~..C C':'CTGTTTGGGGAAACC G W..CnC.:..C CATCAT TAATG T T C:'
AGCCGAT~C.=.vevaAC.'~'nT C~,GTe,TACTCTGCCAV T G.; T G C~,GV.aCTTaG T GGTGG~aT
ATGGAAG ~, ... . . _ .. . ...........FTC~~~,. ,. ~_ _ ~.. ... .... ,.. ,. ..
,."
1'tfVtltZtW L-~i.T~VL.tlil ~.wtSVVSZtiita'..7tli~yL,tinTVt1Vt11Vl'lTVt1
AAGCAATGAA - -- _ _ ..".., ...." .., .......,..,.,.... ~ _ ..._ ,. _ _ _ .. _
.."...... _ .,
'tffl~'lip.~:il~lVtfV'111GiVlaVVI.V.a~illVVilV't'11VW ~.~~L,CAGt~l
SUBSTITUTE SHEET (RULE 26)
CA 02335346 2001-O1-18
WO 00/05360 PCT,'CA99/00656
~8
TA9LE 5 - ZSARA1 Continued
GGCAGACTCTCACCT TGTGTG T GTG ~~AAACGATGATGGCAATTaCCr.G.:.C C ~.=,GGCA
ATTAGTATCCACAAACAACCACGTAAAGTGACCGGGGCCAGCTTCTTTGTC'C=CAGTG
GTGCACTAAAGTCTTCTTCCGGATACCTGGCCAAATCCAGCATAGTAG~.G~:.TGGGGT
AATGGTTCAGATCACCGCAGAGAGCATGGATGCCCTCAGACAGTCCCT';'CGGGAGATG
AAGGATTTCACCATTACATGTGGAAAAGCTGATGCAGAGGAGTCACAGG:..aC~.TGTCT
ATGTCCAGTGGGTGGAGGATGACAAGAACTTTAACAAAGGAGTT':TTAGTCCAATCGA
TGGCAAATCAATGGAGTCTGTGACCAGCGTCAAGATTTTTCATGGCTCAGAATACAAA
GCTAGTGGAAAAATAATTCGCTGGATAGAGGTCTTCTTTCTGGACAATGAGGAGCAAC
AGAGTGGCCTGAGTGACCCTGCTGATCACAGCCGACTCACTGAAAATGTGG.:CAAAGC
ATTCTGTTTAGCGCTTTGCCCACACCTCAAGCTACTGAAGGAAGATGGAATGACCAGG
TTAGGTCTGCGGGTGTCACTGGACTCAGACCAGGTTGGATACCAAGCTGGGAGCAATG
GGCAACTCCTGCCTGCCCGATACACCAATGATTTGGATGGTGCTTTGGTACCAGTGAT
ACACGGGGGCACATGCCAGTTAAGTGAAGGGCCTGTCAGTATGGAGCTGATATTTTAT
ATCCTTGAGAACATCTCCTAGGAAAGACACATGTGTCTCCTCACAAACTGCCATCGCC
CAAACCATTTGCACTTTAACCGCAAAAGATTCATTTTTCTTTTCTTTTGCTAACACTA
GTATTAGGTCAGGGTGCGAGAGGCAGACACCTGAACTCTTAAACC:TCTATGCATTTT
CACAGTAAGGATCAAGCTGCAGCTGGGAATTTCCTGTTACTAATCCAATGTGGGACGT
TAGAAGTGATCGGTGGCACTGACTATCTAGCTGTTCAACCTTCTCTGGCTCCTCTAAG
GACTCTAGTGCCAGGGGGTGAGACATTCAAGTTTAAAACGAAAACTCTAAATACAATC
AGGAATCTCACTCTGACCTCATTTAAATCATCACTGCGACTTTTTTTCCTGCTCGCAT
TCTTTATTTTGCATCTTACTCAAGTTTACATTGTCAAGACCAGCCTAAGCCTTCAGTC
CTTTCTCAATTAAACTACTCGTGCATGGCAAGGAGACTTTCGTTGCACAGCCTGAAAT
ATACCAATCACTTCCCAAACCACAAGCATGAATCCAACGTTTTCCTGACTGGTTGGCT
CTGCTGTGAAAGGGACAGCAATATTATTTTTCTACAGTTGACAAAACTTTTGTCTATG
TCTGTGTCTCTCATGGGGGATTTGTTGCCTGATGGGCAGCCTCCGGAGAGAAGAATTC
CACCCGTGTGTAATATACAGTCTAAGTGTATGGTCTGCTATGTAACACCTGTTGCGCA
GTGCAAATGCACTGACTCTCTGGAAGGCTATAGAGTTTTAAAAACGGTTAGTCTTTTA
AT~~AA.A.AAAA
SUBSTITUTE SHEET (RULE 26)
CA 02335346 2001-O1-18
WO 00/05360 PCT'1CA99100656
59
TABLB 6 ZS~1 - Seauence ID NO_s6
MPKMVIGDTIMAEDSLFNNTGPSEIVCNPSTVESQSLQALDDQSVNIHNEKSVLi~IDG
FSPCSSPKSiINFDCLTMDNEMPLHSQMSVDDNDKETVTISVLPTIIQDTSNVSTDPA
INRPGTKEPHRALKETTSVILPEIKPYSTCAALSFENNNKVPSYQLNNTDLLSVSPW
EACSEQQQKHTSSLHEEKLFEGVSATESFAATAAETVLDNEALRSAEFFDIVVKNFSD
SCVINGDLTKSCGLSQESNEKFCASKEFEGGVDANVLLENACVAYKEAIDLPEENGTN
APMSLYNGCDSYGMKNPAVAQNPKNLPSKEDSVTEEKEIEESKSEYYTGVYEQQREDD
VTERGGLLLNARADQMIQJNLBSLCNQVPSMHGQTSPRRGKIVQSLSVPYGGARTKQPT
~KI~iIPKPLSEMLQSDLIPPNAGCSSKYHI~TDMLNKSNQGDNLISESLREDSAVRS PV
TDANGDFPGEYRGPGSLCLAVSPDSPDNDLLAGQFGVPISKPFTTLGEVAPVWVPDSQ
APNCMKCEARFTFTKRRHBCRACGRVFCAACCSLKCKLQYMDKKEARVCVICHSVLMN
AQAWENMLSASVQSPNPNNPAEYCSTIPPMQQAQASGALSSPPPTVMVPVGVI,I~ipGT
EGSQSKEQRRVWFADGILPNGETADSDNANVTTVAGTLTVSHTNNSTSSESENTSGFC
GSITQVGSAMNLIPEDGLPPILISTGVKGDYAVEERPSQMSVMQQLEEGGPDPLVFVL
NANLLAMVKIVNYVNRKCWCFTTKGMHAVGQAEIVILLQCLPDEKCLPRDLFSHFVEL
YQEAIAGNWGNLGHSFLSQSFLGSIC78GGFLYVAPTYQSLQDLVLPAEPYLFGILIQ
RWETPWAKVFPIRLMLRLGAEYRLYPCPLFSVRYRKPLFGETGHTIINVLADFRNYQY
TLPVVQGLVVDMEVRKTSIKIPSNRYNEMMKANBJKSNEHVLAIGACFNQMADSHLVCV
QNDDGNYQTQAISIHKQPRKVTGASFFVFSGALKSSSGYLARSSIVEDGVMVQITAES
MDALRQSLREMKDFTITCGRADAEESQEHVYVQWVEDDKNFNKGVFSPIDGKSMESVT
SVRIFHGSEYKASGKIIRWIEVFFLDNEEQQSGLSDPADHSRLTENVAKAFCLALCPH
LKLLKEDGMTP.LGLRVSLDSDQVGYQAGSNGQLLPARYTNDLDGALVPVIHGGTCQLS
EGPVSMELI:YILENIS*
SUBSTITUTE SHEET (RULE 26)
CA 02335346 2001-O1-18
WO 00/05360 PCT; C.~99/00656
T71BLE ? aSARA2 Sentience ID NO~?
AGTTTTATTTTCAGAAGACG~~GCATCTTTATTTTAAACATTAAGTTTCACTATGTAG
TAAAACATTACTGTTGTATATACAGTATGTTGTAGACATATAACGTAACTGTTTGCTT
TGTGCTTTCTTTCCTCCTCAGATGAAACTGTCTTTCCAAAGCTGTTAGATGCTAAGTG
GAATCAATTCTTAGAACCACATTCGCATAAAGTCACTGATAAACCAGCTCTTGACAAT
GTCTGTAAATCAATCATTGCTATTGAAGCTCATCTCAAAGTCAGGTCACCCAGCTTGA
CAGCCCTTGCAAGGTCCACATATGTGAATGGAGAAGTAGGTATTGTGACTCCTGAAAT
GCCTAAAATGGTGATAGGAGACACCGATATGGCAGAGGATTCACTTTTTAACACTGGT
CCCTCTGRAATTGTATGCAACTCTATTGTGGAGAGTCAAAGTTTAGAAGTTTTAGATG
ATGTACCAGTGAGTATTAACAATGAA,A,AAAGTGTTCTTCTTGATGATGGATTTTCTCC
GTACAGTAGCCCCAAAAGTGTTCTAAACTCTGCTTGCTTGACCATGAATAACGGAAAG
CCCTCACACGGTCAAAAAATTGTTAATGACCAAGATAAAGAAGCTGTAACAATTTCAG
TCCTTCCAATGATCATACAGGATACTACTAACGTAAGCACAGACCCAGCTTTCAATAA
ATCTGGCACTGAAGAAGCTTATAGTGCATTAAAACAAACCACATCAGTTATTCTGCCT
GAAATAAAGCCTTATTCCATACAGGCTGCCCTTTCATGTGAAAATATCAACAAGATAC
CCAGATGTCAATTAAATAATACAGATCTACTCAGCATTTCACCAGTGGTT'GAAGCATG
TAGTGAGAAGCAGCAAAATCATACAACTTCCTTGCATG1~~AAAAAAACTTGCAGCTGTG
TCTGCAACTGCGTTCTTTCCAGTCACTGCTGCTGRAACTGTACTAGGTAATGAAGCTC
TCCATAGTGCTGATTTTTTTGACATTGTTGTAAAGAACGTTTCTGACTCGTGTGTGTT
TAATGGTGACCTAACTAGAACTAATGGACTCTCACAAGAAAACAATGAAATGTTTTAT
GCAAGTAAAGAGTTGGAAGGAGGGGTAGATGCTAATATCTTATTGGAAGATGCATGCA
TAGCTTATAAAGAAAGAATAGATTTGTCTGAAGAAAATGGAACTAATGCACCAATGTA
TCTGTACAATGGGTGTGATTCCTATGGAATGAAAAACCCTGCTGTACGTCAAAACCCA
AAGAATTTACCATCAAAAGAAGATTCTGTGACAGAAGAAA.AAGAAATTGAAGAAAGCA
AGTCAGAATACTATTCTGGTGTTTATGAACAACAGAAGGAAGATGACATAACTGAGAG
AGGTGGAGTCTTGTTAAATGCCAAGGTTGACCAAATGAAGAACAGTTTGCATAGTCTT
TATAATCCGGTTCCATCCATGCATGGGCAAACCTCACCAAAAAAGGGCAAGATTGTGC
AATCCCTCAGTGTTCCATATGGTGGAGCTCGCCCCAAGCAGCCAACTCATCTCAAACT
CAATATTCCACAGCCATTGTCTGAAATGTTACAGTGTGATCTCATTCCGCCAAATGCT
GGATGCAGCTCTAAA,A.ACAAA.AATGACATGTTAAACAAATCAAATCGGGGGGATAACC
TGATTTCAGAATCACTACGTGAGGAAGTGCACAGCCCTGTTACTGATACAAATGGTGA
AGTCCCTCGAGAAAACAGGGGACCTGGCAGCCTGTGCCTTGCAGTGTC':'CCAGACAGC
CCTGACAATGATCTGCTTGCTGGACAGTTTGGGGTACCCATCTCTAAGCCATTTACTA
CTCTAGGGGATGTGGCTCCAGTCTGGVGTGCCAGATTCCCAAGCr.CCAAACTGCATGAA
GTGCGAGGCCAGATTTACATTTACCAAAAGGAGGCATCACTGCCGAGCTTGTGGAAAG
GTATGTAAAGAAATGTGGTGTTTCATCAGGGCAACAGTAATCACGGCAAATTATTCAT
AACAAAATGTGTTCAGCAGATTCAGTTAAAGTAGACTTATAAGT~ACACAGTAACAAT
TCATCTGCTCAGCCTCATT':'TGAAGTAGATAAAATATATTTTATTAGGAAACTCTGGG
GAGATATAAGGGAAAGCTTGCCTAAAAGTAGATGTTCTGTATATTATTTGGTAGTCAA
AGATGATTTCATGAAAAAAGGTTATTTGTAAAAAGTACAAAATGGG~AGAGACTAGAC
AATAAAAAGTAAGGAGTAAAAAACTAGGTATGTAACGTATATTAAAATAATTTTATGA
TTTTAATATTTACTGCACATT.'TCTACAGTGCAGTGATTTGTATAACCATGCAATTAT
CAAATGCTTAGTGCCTTCACACAAAGTG CCTTTAATAAA.AAT T AT'r"""'ATAAATTATC
ATATTTTCTTTATATGTAGTCATCATCTTTTTTGTCTCATTTC':'~'GGAATCGTTCTAC
TTATGTTCTACTGATATGTT':":'TTACCCGAGACCTATCTTGTCC~'~=AAAGTAATTGG
CTTGTCAACTGGCTGTAGGG;~W,T T'ITCAGAGTTATAGC'~'=AGT:~C'~'GT I'AATGAGCC
ATAGGTTGAAATAGTGC T C'~AGATTTr,CATGTTGTACAACAG T T A:'='"CAATr.TGTGT
AGGGGGGGGG
SUBSTITUTE SHEET (RULE 26)
CA 02335346 2001-O1-18
WO OOI05360 PCTlCA99/00656
61
T~8LB 8 ZSARA2 Seauence ID N0:8
MPRMVIGDTDMAEDSLFNTGPSEIVCNSIVESQSLEVLDDVPVSINNEKSL4LDDGFS
pYSSPRSVLNSACLTMNNGRPSHGQRIVNDQDREAVTISVLPMIIQDTTNVSTDPAFN
KSGTEEAYSALRQTTSVILPEIRPYSIQAALSCENINRIPRCQLNNTDLLSISPVVEA
CSERQQNHTTSI,HEKRLAAVSATAFFPVTAAETVLGrtEALHSADFFDIWF~TVSDSCV
FNGDLTRTNGLSQENNEMFYASRELEGGVDANILLEDACIAYRERIDLSEENGTNAPM
YLYNGCDSYGMRNPAVRQNPRNLPSREDSVTEEREIEESKSEYYSGVYEQQKEDDITE
RGGVLLNARVDQMRNSLHSLYNPVPSMHGQTSPKRGRIVQSLSVPYGGARPKQPTFiLK
LNIPQPLSEMLQCDLIpPNAGCSSRN~Q~TDMLNKSNRGDNLISESLREEVHSPVTDTNG
EVPRENRGPGSLCLAVSPDSPDNDLLAGQFGVPISRPFTTLGDVAPVWVPDSQAPNCM
KCEARFTFT~GtRHIiCRACGKVCREMWCFIRATVITANYS
SUBSTITUTE SHEET (RULE 26)
CA 02335346 2001-O1-18
WO 00/05360 PCTlCA99/00656
62
TABLE 9
hSARA AE IKR'I~YS~O~AYE E CG NYlVYIYWMN OK11T~NOlODBNh~hS~lC .~
XSARA ~rK TI AED fNNfTIGPSE'IV~NPSTlI~B~-. p .....vh~yNlEKSVtJADGfSIe35r - fmn
70
hSAfv1 ac L ENROTDOfBiSINES D NiE ,,l~ t KT v ITSI StASY(C7S S t~t00G51 K013~5
tS~
XSARA cO~ YO~...............1LN= Lf~N .. Tiy~~TLSNVS~ AI~N~tlr. ~Kfr~o ttt
hSARA 1 St DI ItS TOGP.f v K(~~v~E~KISSIR~INSF~~ILA1KA NTS~LN~~FE'V6 111
XSApA t c E T~ Y I t~P E I ~I V 3 T~A~ F ELN~,~,~,,~, KL~'I 3 v NN - N p . v C
3 EI00
hSARA tttKP011PNGIGRNNOCERCSOCtviHEVp~ENrEwE~tN~ESE30D~t~K~KtN~DSOV C
XSARA aTES-~-~~-~---..............cA AA VtON IL AE
FDI~VCLSL'.iSOSC~UItI~~L'.'O~TKIS ~C
hSARA KOIS~~TNGOS~CVGLA, AGt ~~f~~FSTVI~~NV~ GCDSVGII ~~cv K LPS 791
XSARA NEK aSKEfC4J1---.. .vpANVt~E ACVA K 1 LPEEN~~SL~~C~6w3LGL~
OCDfYDItKIrL6JIAl.~AON KNLPf107t1
XSARA ~DfVTEIKIIEEfKf ITGV~E 0~ O~Y EIS LLLNATII~p~l~1''NSL~S
VIStINKtOIfIKIKGKSV tSVPV~711
hSARA A OPSNLIL0,1~KPLiD~ ~L N~ f A S N~,. N011GKA L E SA1 YCSP.~tGNISNV NGE
LES EAE SS7
XSARA ~~ O~TN~KLN71K~LSE~S~t I~r-AD)ct~ 81D11LNK! ONLI~ESLRE061AVR3VVT
ANOOIFPGEQIIGI a71
~fw111f 1
hSARA ~RP LAtAPDiPOND_R GOfGI AR
~._LGEYAPVWVPDi0A1NC11_KCEARFTFTKRRNNCRACOKYFGASGGi 117
XSARA '~'6Ct) - l tAVSIDfIDNO L GOFOVP t S'CP.F:TT-
LOEVA~VWV1DlOA~NCitKG~~RF:TFTKIIRNNCRACOKVFCAACCSLI SSE
t
hSARA C YOR YCYJLN VlIWAOAWENYYiA _NPNN1A
.~_.ifl.~lLpaAOAG_GALSSPIPTVIIV~YOYiKN 117
XSARA CK iIOKKEARVCVtCNi iNAOAwENtlli~f
f~N1NN~~.CfTTPP400A0Ai0ALiiIPPTVYVIVOVIKN i7t
~ P~2 ___-
hSARA -OppVWPA OtLPf~GE DAAKITLA ._ IVKIV~T~LPAETOI SGiI-TOV SIY~T87
XSARA T - LOpRVWFADGfLTNOE DSD-- QTLT f TNNSTS -ENT CGfTTOV -~~ TQS
hSARA NLJPEOGLIII.LIf
OV~KOD_Y;AVEEK1f01iY110OLE0a010V,LVFV,LNANLLSWKI.VNVVNRKCWCFTTKGIINqYG 877
><SARA NL~IPED01PPIC'IfTOVKGDYAVEE11~f01tfV1100LEE00IDPLVPVLNANLfLAWKI-
VN1'VNRKCWCF7TKOIINAVG T85
hSARA SELVI:LLOCL10EKCLPK01 N iVOLT.RD LAONYYS LGNfF OSFLOiKENGGf.LYVT
vOSLODLYLP.T ELF fi3
:SARA aEIVICLOCLPDEKCLIRDtilS ' "~FVEL'fOE~IIAONVVIG LONf Of FL GSKONGGFITVaP
YOSLDDLVLPA ~Yli 88f
hSARA li'IOKriE.TPWAKVfPIRLYLRLGAEVRLT CPLFSVpFRKPL O
aNTt:AtNtLADfRNVOVTLPVVOOLVVD11EYRK 1017
'.-_.
XSARA '7lCtCKWET~WAKVFPStI~yLR~'GAEY!L'YPCPLFSV11y11K1L~FOETONT~
IGIV~AOFRN110YTL1VVOGL:VVDHEVRKTIItf
hSARA IKIPiNp~NEIIYK~YWKiNENVL ACfNE DSNLVCYONOOGNVOTOA15.1. N
P_RKVTGAiFFVFSGALKSSiO 1117
XSARA I KIPfNp~NEAIYItAINKfNENVLAIWACPNOa DSNLVCVDNDDONItOTOAISt
NKGPRKVTOAfFFVFSOALKSSiOY~ tD25
hSARA AKfi_1 VE_DOV11V01 TAE N S ROALREWLDFTI TGGKADAEE EN t I~ DWV ODD_K S
OY,Y P i OGK511ET l 1197
XSARA -AKfl7VEDGVYVO(TAES~DA~ROSLREtucDFTITCOKADAEE EHV ~bwyEDDKNicNKGVIF
PIOGKSAESV ~1 Ittlf
hSARA IfJtGiEYK KYIIR T V _LENODON~NC
iDPAONSRI:TENYAK~fGLALCPNLKLLKEOGN1TKLGLR1ITLOSDOV.G 1177
XSARA IfNafEYK KIlR I YFFI:DNEE001SG-fDPAONSRLTENVAKAiCLALC1-
NLKLLKEOGAtTRLGLRV3LDSOOVO ttlS
hSARA AGtN ILPSOY OLDS ~/~L~.V~~(~ KtliGOAGOLiEOPYV ELI_fYILENI . tJZ7
XSARA b~OfN L 11R.~ DLO Ia=r/t'VIiNGGT=OLSEOPVS ELI FYILENI . t37S
_ TABLE 10
hSARA Sf7~ ~~~~ ~ 5~. w .~.tD./'nl. -E~y I~SV 655
XSARA St0 GE ~ ~P PNG11KCEIAR ~ KRpNNCRAGOKVFC A~ SiLkCKLCIIYaDKK.EhRVCI'lI~SV
K1AA0305 777~'- GO~~~T-T~~ APNG11f7tONK IIrr~
KRRNNCRACOKVFCCLfiC(:FKCKLDKl:EKF E~ VCNV TI 800
FGD7 nO~OR~1~~T I ~(~rrT~IAT~C01E ~~K11RNNCKACGNV GkCS~~AALNkhJNNR.~~V,C%'0 A
485
lin -tS~I'~iiA1 ~. -'.~E Rp~'VG~F ~',NiTTRKNNG1ACG01~GKC55R17~t'~. p'fG ~
EK~JPVCEP Y 0 ytg
Hrs-2 t53AAERAP~ D. . ~AEE~HPCtavOFE GNP tRKHNCRACGOIFCGkCS5Kn6Gl! PK ~G ~
EKE4 VCEP Y 0
EEA~13t1TOAtN K E~T~IEF~OFTC~KuIF4~-=T?.rpRNNCR~GG11FC~EtSAKNN~'~=SSK~-r.-
.'v~VCDA FND~t408
GTLS~.6.....rD.W...... -.C..C...a=.....~~~.~~.~........-..-..
SUBSTITUTE SHEET (RULE 26)