Note: Descriptions are shown in the official language in which they were submitted.
CA 02355198 2001-07-18
73529-137D
- 1 -
ALS3 PROMOTER IN TRANSFORMED PLANTS
This application is a divisional of Canadian Patent
application No. 2,216,244 stemming from PCT application
PCT/US96/03635 filed on March 22, 1996.
BACKGROUND OF THE INVENTION
I. Field of the Invention
The present invention relates to a novel ALS3
promoter which generally drives constitutive and generally non-
tissue-preferred expression of operably linked foreign genes in
transformed plants. In particular, this invention is directed
to, DNA constructs in which a Brassica ALS3 promoter is operably
linked to a foreign structural gene, and to using the DNA
construct to produce, in a transformed plant, a protein which
is encoded by the structural gene. The Brassica ALS3 promoter
is used to direct expression of agronomically important genes
and selectable marker genes.
II. Background
Acetolactate synthase (ALS), which is also known as
acetohydroxy acid synthase (AHAS), catalyses the first step in
the biosynthesis of the branched chain amino acids leucine,
isoleucine and valine. It has also been shown to be the site
of action of sulfonylurea and imidazolinone based herbicides.
See, for example, Chaleff, R.S. and C.J. Mauvais, Science
224:1443 (1984) and Shaver et al., Plant Physiol. 76:545
(1984). A number of different ALS genes from Brassica napus
have been cloned and characterized. See, for example Wiersma
et al., Mol. Gen. Genetics 219:413 (1989) and Rutledge et al.,
loc. cit. 229:31 (1991).
Rutledge et a1. (1991) reported that the B. napus
rapeseed cultivar Topas contains an ALS multigene family
CA 02355198 2001-07-18
73529-137D
- la -
comprised of five genes. DNA sequence analysis of the
structural genes revealed that the ALS1 and ALS3 genes shared
extensive sequence homology. In contrast, the ALS2 gene has
diverged significantly from the ALS1 and ALS3 genes and has
unique features in the coding region of the mature polypeptide,
transit peptide and upstream non-coding region. The ALS2 gene
therefore may encode a
CA 02355198 2001-07-18
~, _ i6I30530 PCT/US96I036.,_
_ 2 _
polypeptide with a distinct function from that of ALS1
and ALS3. The ALSO and ALSS genes have interrupted
coding regions and therefore may be defective.
Experiments conducted with the promoter of the
Arabidopsis thaliana ALS gene revealed that the A.
thaliana ALS promoter is significantly less effective in
driving gene expression than the CaMV 355 promoter.
Odell et al., Plant Physiol. 94 4 :1647-1654 (1990)
replaced the A. thaliana ALS promoter with the CaMV 35S
promoter and observed a 25-fold increase in the level of
ALS mRNA accompanied by a 2-fold increase in ALS enzyme
level and a 3-fold increase in sulfonylurea tolerance.
These observations indicate that the ALS gene is
regulated post-transcriptionally and that the A. thaliana
ALS promoter is significantly less effective in driving
gene expression than the Ca~'~IV 35S promoter.
The number of isolated and characterized constitutive
generally non-tissue-preferred plant promoters available
for expression of foreign proteins in transgenic plants
is very limited. Well known examples of promoters with
constitutive and tissu= generated expression patterns
include those associated with the CaMV 3SS, Agrobacterium
nopaline synthase, and maize ubiouitin genes. See Odell
et al., Plant Mol. Biol. 10 3 :263-272 (1988), Herrera-
Estrella et al., Nature 303:209-213 (1983) and Fox et
al., Va. J. Sci. 43 2 :287 (1992).
There is a critical need for a broader repertoire of
strong constitutive anti generally non-tissue-preferred
plant promoters. A broader array of constitutive and
generally non-tissue-preferred plant promoters that are
expressed at high levels, that is, that drive expression
of operably linked genes at a level comparable to the
CaMV 35S promoter, would allow the genetic engineer to
analyze the relative strengths of the available promoters
and select promoters that provide the required level of
expression of foreign genes in transformed plants. A
CA 02355198 2001-07-18
. .. .r 96130530 PCT/U596103~_.,
- 3 -
selected promoter might provide optimum levels of
expression for the first gene but may be either too
strong or too weak for use in driving the expression of
a second gene. Consequently, additional constitutive and
tissue general promoters are needed to optimize foreign
gene expression in plants.
. There is also _a need for additional strong
constitutive and generally non-tissue preferred promoters
for construction of plants transformed with multiple
foreign genes. Numerous difficulties have arisen when
two or more different genes are introduced into a plant
wherein each of the genes are operably linked to the same
. or similar promoters. Some of these difficulties include
~(1) gene inactivation; (2) recombination as a result of
pairing along homologous regions within the nucleotide
sequence of the promoter leading to cross-over events and
loss of the intervening region prior, or subsequent to,
integration; and (3) competition among different copies
of the same promoter region for binding of promoter-
specific transcription factors or other regulatory DNA-
binding proteins. A need therefore exists for a broader
repertoire of strongly constitutive and tissue general
promoters to be used for expression of foreign genes in
transformed plants.
SUMMARY OF THE INVENTION
Accordingly, it is an object of the present invention
to provide a strong, constitutive promoter which can
effect high level, generally non-tissue-preferred
expression of an operably linked, foreign gene in
transformed plants.
It is another object of the present invention to add
to the limited repertoire of generally non-tissue-
preferred promoters available for the transformation of
plants with multiple genes.
In achieving these and other objects, there has been
provided, in accordance with on=_ aspect of the present
CA 02355198 2001-07-18
~6I30530 PC'TlUS95/036.._
- d -
invention, an isolated DNA molecule comprising a promoter
operably linked to a foreign structural gene, wherein
said promoter comprises a nucleotide sequence
corresponding to the sequence of a polynucleotide from
the group consisting of (1) a XbaI/Ncol fragment 5-prime
to the Brassica napus ALS3 structural gene or (2) a
nucleotide sequence .that has substantial sequence
similarity with said XbaI/NcoI fragment. Other objects
of the present invention include providing an isolated
DNA molecule wherein the promoter is operably linked to
an agronomically important gene or a selectable marker
gene, an isolated DNA that is part of an expression
vector and an expression vector carrying the isolated DNA
molecule that is present in a transformed host.
It is another object of the present invention to
provide an isolated DNA molecule comprising a promoter
operably linked to a foreign structural gene, wherein
said promoter comprises a nucleotide sequence
corresponding to the sequence of a polynucleotide from
the group consisting of (1) SEQ ID NO: 1 or (2) a
nucleotide sequence that has substantial sequence
similarity with S~Q ID NO: 1. Other objects of the
present invention include providing an isolated~DNA
molecule wherein the proTOter is operably linked to an
agronomically important gene or a selectable marker gene,
an isolated DNA that is part of an expression vector and
an expression vector carrying the isolated DNA molecule
that is present in a tra.~.sform~d host.
Another object of the present invention is to provide
a method of using an ALS3 promoter to produce a foreign
protein in a transformed host plant, comprising the steps
of (1) constructing an expression vector comprising a
promoter operably linked to a foreign structural gene,
wherein the promoter comprises a nucleotide sequence
corresponding to the sequence of a polynucleotide from
the group consisting of (a) a XhaI/NcoI fragment 5-prime
CA 02355198 2001-07-18
. J 96130530 PCTIUS96IO~~~S
- 5 -
to the Brassica napes ALS3 structural gene or (b) a
nucleotide sequence that has substantial sequence
similarity with said XbaI/NcoI fragment; and (2)
transforming a host.
Yet another object of the present invention is to
provide a method of using an ALS3 promoter to produce a
foreign protein in a transformed host plant, comprising
the steps of (1) constructing an expression vector
comprising a promoter operably linked to a foreign
structural gene, wherein said promoter comprises a
nucleotide sequence corresponding to the sequence of a
polynucleotide from the group consisting of (a) SEQ ID
NO: 1 or (b) a nucleotide sequence that has substantial
sequence similarity with SEQ ID NO: 1; and (2)
transforming a host.
Other objects, features and advantages of the present
invention will become apparent from the following
detailed description. It should be understood, however,
that the detailed description and the specific examples,
while indicating preferred embodiments of the invention,
are given by way o~ illustration only, since various
changes and modifications within the spirit and scope of
the invention will become apparent to those skilled in
the art from this detailed description.
BRIEF DESCRIPTION OF THE DRAWINGS
Figure 1 presents the nucleotide sequence (SEQ ID N0:
1] of a 1063 by XbaI/NcoI fragment that comprises the
promoter ~-prime to the B. napes ALS3 wild type
structural gene. The XbaI and NcoI restriction sites are
underlined. The ATG start codon is found within the NcoI
restriction site.
Figure 2 presents a restriction man of a 1063 by
XbaI/NcoI fragment which comprises the B. napes ALS3 wild
type structural gene.
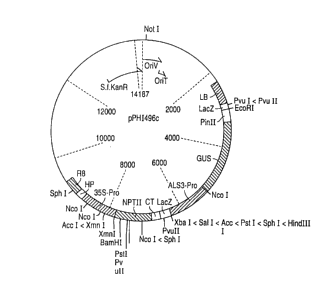
Figure 3 presents a man of pPHI4960, a binary vector
containing the B. napes ALS3 promoter (ALS3-Pro) driving
CA 02355198 2001-07-18
~. _ 16130530 PCT/US96I036~_
uidA (GUS) gene expression. Plasmid pPHI4960 was used to
transform canola and sunflower. LB and RB represent th=_
left and right borders, respectively, of the
Agrobacterium Ti plasmid T-DNA region. The region
between the left and right borders also includes a
CaMV 35S promoter (35S-Pro) driving expression of the
NPTII gene for kanamycin selection of transformed- plarits.
Figure 4 presents a map.of pPHI6333 which is a binary
vector containing a cassette comprising the B. napus A.LS3
promoter (BNALS3 PROM), NPTII gene and the 3-prime
terminator sequence from the potato proteinase inhibitor
gene (PINII). Plasmid oPHI6333 was used to transform
sunflower to ascertain the suitability of this promote_~
for driving selectable marker expression. LB and R3
represent the left and right borders, respectively, o
the Agrobacterium Ti plasmid T-DNA region.
Figure S presents a man of plasmid pPHI3042 whic:~
contains the NPTII and GUS structural genes operably
linked to the CaMV 3SS promoter.
Figure 6 presents a map o' plasmid pPHI5765 whic
contains the NPTII structural gene operably linked to the
CaMV 35S promoter (3SSPR0) and the potato proteinase
inhibiter gene terminator sequence (PINII).
Figure 7 is a histogram showing GUS activity in plan=
2S seedlings transformed with expression cassettes in whic:~
the GUS gene is expressed from either the CaMV 35S or
ALS3 promoters. GUS activity was measured in cotyledon,
hypocotyl, meristem and root tissues.
Figure 8 is a histog=am showing GUS activity in
vegetative stage plants transformed with expressio:~
cassettes in which the GUS gene is expressed from either
the CaMV 35S or ALS3 promoters. GUS activity was
measured in leaf, petiole, meristem, stem and root
tissues.
Figure 9 is a histogram showing GUS activity in
flowering stage plants transformed with expression.
CA 02355198 2001-07-18
.. ~ 96/30530 PCT/US96I036~~
cassettes in which the GUS gene is expressed from either
the CaMV 35S or ALS3 promoters. GUS activity was
measured in flower, leaf stem and root tissues.
Figure 10 is a histogram showing GUS activity in
mature plants transformed with expression cassettes in
which the GUS gene is expressed from either the CaMV 35S
or ALS3 promoters. GUS activity was measured in leaf,
stem, pod and seed tissues..
DETAILED DESCRIPTION
I. Definitions
In the description that follows, a number of terms
are used extensively. The following definitions a=a
provided to facilitate understanding of the invention.
A structural gene is a DNA sequence that is
transcribed into messenger RNA (mRNA) which is then
translated into a seauence of amino acids characteristic
of a specific polypeptide.
A promoter is a DNA sequence that directs the
transcription of a structural gene. Typically, a
promoter is located in the S' region of a gene, proximal
to the transcriptional start site of a structural gene.
If a promoter is an inducible promoter, then the rate of
transcription incr eases in response to an inducing agent .
For example, a promoter may be regulated in a tissue-
2S specific manner such that it is only active in
transcribing the associated coding region in a specific
tissue types) such as leaves, roots or meristem.
In contrast, the rate of transcription is not
generally regulated by an inducing agent it the promoter
is a constitutive promoter. The promoter may be tissue
general, also known as non-tissue-preferred, such that i t
is active in transcribing th=_ associated coding region i n
a variety of different tissue twes.
A core promoter contains essential nucleotide
3S sequences for promoter function, including the TATA box
CA 02355198 2001-07-18
~ . .. 96130530 PCTlUS96/036_
- 3 -
and start of transcription. By this definition, a cor=_
promoter may or may not have detectable activity in the
absence of specific sequences that may enhance the
activity.
S An isolated DNA molecule is a~~fragment of DNA that
is not integrated in the genomic DNA of an organism. For
example, the promoter of the ALS3 gene is a DNA fragment
that has been separated from the genomic DNA of Brassica
napes.
Complementary DNA (cDNA) is a single-stranded DNA
molecule that is formed from an mRNA template by the
enzyme reverse transcriptase. Typically, a primer
complementary to portions of mRNA is employed for the
initiation of reverse transcription. Those skilled in
the art also use the term "cDNA" to refer to a double-
stranded DNA molecule consisting of such a single-
stranded DNA molecule and its complementary DNA strand.
To operably link one nucleotide sequence to another
refers to joining two heterologous DNA fragments to
produce a chimeric DNA construct that has biological
activity. For exampl=, an isolated DNA fragme::t
comprising a promoter from a first gene, such as the ALS3
gene, is operably linked to an isolated DNA fragment
comprising the structural gene from a second heterologous
gene. The resulting chimeric DNA construct is functional
when the AL3 promoter is shown to initiate transcription
of the heterologous structural gene.
The term expression =e~ers to the biosynthesis of a
gene product. For example, in the case of a structural
gene, expressior.involv~s transcription of the structural
gene into mRNA and the translation of mRNA into one o_~
more polypeptides.
A cloniny vector is a DNA molecule, such as a
plasmid, cosmid, or bacteriophage, that has the
capability of reel icati ng autonomously in a host cell .
Cloning vectors typically contain one or a small number
CA 02355198 2001-07-18
~ 96!30530 PCTlUS96103~__
- 9 -
of restriction endonuclease recognition sites at which
foreign DNA sequences can be inserted in a determinable
fashion without loss of an essential biological function
of the vector, as well as a marker gene that is suitable
S for use in the identification and selection of cells
transformed with the cloning vector. Marker genes
typically include genes. that provide tetracycline
resistance or ampicillin resistance.
An expression vector is a DNA molecule comprising a
gene that is expressed in a host cell. Typically, gene
expression is placed under the control of certain
regulatory elements, including constitutive or inducible
. promoters, tissue-specific regulatory elements, and
enhancers. Such a gene is said to be "operably linked
to" the regulatory elements.
A foreign gene refers in the present description to
a DNA sequence that is operably linked to at least one
heterologous regulatory element. For example, any gene
other than the ALS3 structural gene is considered to be
a foreign gene ii the expression of that gene is
controlled by the ALS3 promoter.
A recombinant host may be any prokaryotic or
eukaryotic cell that contains either a cloning vector o.
expression vector. This term also includes those
prokaryotic or eukaryotic cells that have been
genetically engineered to contain the cloned genes) 'n
the chromosome or genome of the host cell.
A transQenic plant is a plant having one or mora
plant cells that contai-~ an expression vector.
In eukaryotes, RNA pclymerase IT catalyzes t:~:e
transcription of a structural gene to produce mRNA. A
DNA molecule can b= designed to contain an RNA polymerase
II template in wi:ich the RNA transcript has a sequence
that is complementary tc that of a specific mRNA. The
RNA transcript is termed an antisense RNA and a DNA
sequence that encodes the antisense RNA is termed an
CA 02355198 2001-07-18
96130530 PCTlUS96I0~.
- 10 -
antisense ctene. Antisense RNA molecules are capable of
binding to mRNA molecules, resulting in an inhibition of
mRNA translation.
A first nucleotide sequence has substantial sevuence
S similarity to the nucleotide sequence of Figure 1 [SEQ.
ID NO: 1] if the former sequence share a similarity of at
. least 65% with the Figure 1 sequence and is a
constitutive promoter active in directing the
transcription of an operably linked foreign structural
gene in plants. Sequence similarity determinations can
be performed, for example, using the FASTA program
(Genetics Computer Group; Madison, WI). Alternatively,
sequence similarity determinations can be performed using
BLASTP (Basic Local Alignment Search Tool) of the
1S Experimental GENIFO~ BLAST Network Service. See Altschul
et al., J. Mol. Biol. 215:403 (1990). Also,. see
Pasternak et al., "Sequence Similarity Searches, Multiple
Sequence Alignments, and Molecular Tree Building," in
METHODS IN PLP.NT MOLECULAR BIOLOGY AND BIOTECHNOLOGY,
Glick et aI. (eds.), pages 251-20'7 (CRC Press 1993).
Promoter activity o~ the isolated nucleotide sea~uence can
be assayed by means of fusing the nucleotide sequence to
a coding region of a foreign reporter gene. Promoter
activity is measured by assaying reporter expression.
See, for example, An et al., "Techniques for Isolating
and Characterizing Transcription Promoters, Enhancers,
and Terminators," in METHODS IN PLANT MOLECULAR BIOLOGY
AND BIOTECHNOLOGY, Glick et al. (eds.), pages 15S-155
(CRC Press, 1993).
II. Cloning of ALS3 Promoters
A 1063 by Xbal/NcoT_ Fragment comprising the promoter
region of the ALS3 gene ~=om wi ld type Brassica napes was
cloned into pGEMS (Promega Corporation, Madison, WI) and
characterized. The nucleotid=_ sequence of the 1063 b~
Xbal/NcoI fragment was determined by the dideoxy
sequencing protocol. Sanger et ai., Proc. Nat'1 Acad.
CA 02355198 2001-07-18
.96/30530 PCT/US96103.
- II -
Sci. USA, 74:5463 (1977). The nucleotide sequence of the
1063 by XbaI/NcoI fragment is shown in Figure 1 [SEQ ID
NO: 1J and a detailed restriction map of this same
fragment is shown in Figure 2.
Other ALS3 promoters having substantial sequence
similarity with the nucleotide sequence shown in Figure
1 can be cloned by conventional methods.
Oligonucleotides of defined sequence are chemically
synthesized. Itakura et al., Annu. Rev. Biochem. 53:323
(1984). Numerous automated and commercially available
DNA synthesizers are currently available. The probe can
be a single and relatively short oligonucleotide of
defined sequence, pools of short olgonucleotides whose
sequences are highly degenerate or pools of long
oligonucleotides of lesser degeneracy. Sambrook et al.,
Molecular Cloning: A Laooratory Manual, 2nd ed. (Cold
Spring Harbor Press, Cold Spring Harbor, N.Y. 1989).
The oligonucleotid~ hybridization probes based on S3Q
ID NO: 1 are labeled, fo. example radio labeled, by
conventional methods and used to detect related
nucleotides sequences in Brassica genomic libraries by
means of DNA hybridiza~ion. See, for example, Sambroo'.<
supra.
A plant genomic DNA library can be prepared by mews
well-known in the art. See, for example, Slightom et a1.
"Construction of ~ Clone Banks," in METHODS IN PLA_~i
MOLECULAR BIOLOGY AND 3IOTECHNOLOGY, Glick et a1 . (eds. ) ,
pages 121-146 (CRC Press, 1993). Genomic DNA can be
isolated from Hrassica tissue, for example, by lysi::g
plant tissue with the d~t~rgent Sarkosyl, digesting t~~
lysate with proteinase K, clearing insoluble debris from
the lysate by centrifugation, precipitating nucleic acid
from the lysate using isoDropanol, and purifying
resuspended DNA on a cesium chloride density gradient.
3S Ausubel at a1. (eds.). CURRENT PROTOCOLS IN MOLECUI~.3~
BIOLOGY, pages 2.3.1-2.3.3. (1990),
CA 02355198 2001-07-18
.. ~ 96/30530 PCTIUS961036_
DNA fragments that a=a suitable for the production
of a genomic library can be obtained by the random
shearing of genomic DNA or by the partial digestion of
genomic DNA with restriction endonucleases. See, for
example, Ausubel et al.. supra, at pages 5.3.2-5.4.4, and
Slightom et al., supra.
Genomic DNA fragments can be inserted into a vector,
such as a bacteriophage or~cosmid vector, in accordance
with conventional techniques, such as the use of
restriction enzyme digestion to provide appropriate
termini, the use of alkaline phosphatase treatment to
avoid undesirable joining o. DNA molecules, and ligation
. with appropriate ligases. Techniques for such
manipulation are disclosed by Slightom et al., supra, and
are well-known in the art. Also see Ausubel et al.,
supra, at pages 3Ø5-3.17.5.
A library contain_ng genomic clones is screened with
DNA hybridization probes based on the nucleotides
sequence of the ALS3 promoter shown in Figure 1 (SEQ ID
NO: 1] using standard 'echniques. See, for example,
Ausubel et al., supra, at pages 6Ø3-6.6.1; Slightom et
al., supra.
III. Characterization of ALS3 Promoters
Genomic clones can b~ analyzed using a variety of
techniques such as rest_iction analysis, Southern
analysis, primer extension analysis, and DNA sequence
analysis. Primer extension analysis or S1 nuclease
protection analysi s, nor ~xa«,pl ~, can be used to localize
the putative start site oz transcription of the cloned
gene. Ausubel et al., supra, at pages 4.8.1-4.8.5;
Walmsley et al., "Quantitative and Qualitative Analysis
of Exogenous Gene expression by the S1 Nuclease
Protection Assay," in MST:-iODS IN MOLECULAR BIOLOGY, VOL.
7: GENE TRANSFER A:VD EXPRESSION PROTOCOLS, Murray (ed.),
pages 271-281 (Humana ~=ess Inc. 1991). Structural
CA 02355198 2001-07-18
J 96130530 PCT/U596/0~,,_~
analysis can be combined with functional analysis for a
complete characterization of the promoter region.
The general approach of such functional analysis
involves subcloning fragments of the genomic clone into
an expression vector which contains a reporter gene,
introducing the expression vector into various plant
tissues, and assaying_the tissue to detect the transient
expression of the reporter gene. The presence of a
constitutive, tissue-general promoter is verified by the
observation of reporter gene expression in diverse plant
tissues including roots, stems or leaves.
Methods for generating fragments of a genomic clone
. are well-known. Preferably, enzymatic digestion is used
to form nested deletions of genomic DNA fragments. See,
for example, Ausubel et al., supra, at pages 7.2.1-
7.2.20; An et al., supra.
Alternatively, DNA tha~ resides "upstream," or S-
ward, of the transcriptional start site can be tested by
subcloning a DNA fragment that contains the upstream
region, digesting the DNA i=agmer.t in either the 5' to 3'
direction or in t:-~e 3' to 5' direction to produce nested
deletions, and subcloning the small fragments into
expression vectors for c=ansient expression.
The selection of an appropriate expression vector
will depend upon the method of introducing the expression
vector into host ells. Typically, an expression vector
contains: (1) prokaryotic DNA elements coding for a
bacterial replication crigin and an antibiotic resistance
marker to provide for the growth and selection of the
expression vector in the bacterial host; (2) eukaryotic
DNA elements that control initiation of transcription,
such as a promoter; (3> DNA elements that control the
processing of transcrip t, suc:~. as a transcription
termination/polyadenylation sequence; and (4) a reporter
gene that is operably linked to the DNA elements that
control transcription. in_=iation. Useful reporter genes
CA 02355198 2001-07-18
'. .. 96/30530 PCTIUS96/036__
- I4 -
include a-glucuronidase,~3-galactosidase,chloramphenicol
acetyl transferase, luciferase, and the like.
Preferably, the reporter gene is either the ~i-
glucuronidase (GUS) gene or the luciferase gene. See,
for example, Jefferson et al., Plant Molecular Biology
Reporter 5 4 :387 (1987). General descriptions of plant
expression vectors and reporter genes can be found in
Gruber et al., "Vectors for Plant Transformation," in
METHODS IN PLANT MOLECULAR BIOLOGY AND BIOTECHNOLOGY,
Glick et a1. (eds.), pages 89-119 (CRC Press, 1993).
Moreover, GUS expression vectors and GUS gene cassettes
are available from Clontech Laboratories, Inc. (Palo
Alto, CA), whip luciferase expression vectors and
luciferase gene cassettes are available from Promega
Corporation (Madison, P1I) .
Expression vectors containing test genomic fragments
can be introduced into protoplasts, or into intact
tissues or isolated cells. Preferably, expression
vectors are introduced into intact tissues. General
methods of culturing plant tissues are provided, for
example, by Miki et al., "Procedures for Introducing
Foreign DNA into ?lants," in METHODS IN PLANT MOLECUI:A.R
BIOLOGY AND BIOT~CHNOLOG'!, Glick et a1 . (eds. ) , pages 57-
88 (CRC Press, 1993). Methods of introducing expression
vectors into plant tissue include the direct infection or
co-cultivation o~ plan= tissue with Agrobacterium
tumefaciens. Horsch et al., Science 227:1229 (1985).
Descriptions of Agrobact~=ium vector systems and methods
for Agrobacterium-mediated gene transfer are provided by
Gruber et al., supra, and Miki et al., supra. Methods of
introducing expression vectors into plant tissue also
include direct gene Transfer methods such as
microprojectile-mediated delivery, DNA injection.,
electroporation, and the like. See, for example, Gruber
et al., supra; Mik= et al., supra.
CA 02355198 2001-07-18
~ 96130530 pCT/US96/0~
_ 15 _
The above-described methods have been used to
identify and characterize the Brassica ALS3 promoter that
is constitutively expressed in transformed plants. In
particular, the ALS3 constitutive and generally non-
tissue-preferred promoter was found to reside within a
1063 by DNA fragment shown in Figure 1 (SEQ ID NO: 1).
Thus, the present invention encompasses a DNA molecule
having a nucleotide sequence of SEQ ID NO: 1 and having
the function of a constitutive plant promoter.
Variants of the 1063 by generally constitutive and
non-tissue-preferred promoter can be produced by
deleting, adding and/or substituting nucleotides for the
nucleotides recited in SEQ ID~NO: 1. Such variants can
be obtained, for example, by oligonucleotide-directed
mutagenesis, linker-scanning mutagenesis, mutagenesis
using the polymerise chain reaction, and the like.
Ausubel et al., supra, at pages 8Ø3-8.5.9. Also see
generally, McPherson (ed.), DIpECTED MUTAGENESIS: A
PRACTICAL APPROAC:~, IRL PreSS (1991). Thus, the present
invention also encompasses DNA molecules comprising
nucleotide sequences that have substantial sequence
similarity with SEQ ID NO: i and function as a tissue-
general promoter.
Moreover, additional deletion analyses are performed
to further localize the core promoter region within the
1063 by promoter. Thus, the present invention also
encompasses fragments oT the DNA molecule having
nucleotide sequence of SEQ ID NO: ?, as long as the DNA
fragments function as a constitutive promoter.
IV. Vector Construction
The 1063 by Xbal/NcoI fragment shown in Figure 1 [S?Q
ID NO: 1] was fused to a GUS r~po~ter gene cassette which
includes the 3-prim. terminator sequence from the potato
proteinase inhibitor gene (PINIi). The ALS3 promoter was
cloned as an XbaI/EcoRI fragment into the corresponding
sites of the binary vector pP:T.I1741 to create plasmid
CA 02355198 2001-07-18
95/30530 PCTIL1S961036_ _
_ 1' _
pPHI4960 (Figure 3). Plasmid pPHI1741 differs from
plasmid pBI101.1, taught by Jefferson et al., supra, by
having the CaMV 35S rather than the nopaline synthase 5
prime and 3-prime regulatory sequences driving the NPTII
selectable marker gene.
The vector pPHI6333 was constructed by replacing the
region between the.PmeI and EcoRI sites of pPHI1741 with
a cassette containing the ALS3 promoter driving an NPTII
selectable marker gene and the PINII 3' terminator
sequence (Figure 4).
v. Agronomic Genes and Selectable Marker Genes for
Brassica Transformation
By means of the present invention, agronomic genes
' 'and selectable marker genes can be operably linked to the
ALS3 promoter and constitutively expressed in transformed
plants. More particularly, Brassica can be genetically
engineered to express various Dhenotypes of agronomic
interest. The genes implicated in this regard include,
but are not limit'd to, those categorized below.
1. Genes That Confer Resistance To Pests or Disease And
That Encode:
(A) Plant dis=_ase resistance genes. Plant defenses
are oLt~n activated by specific interaction
between the o=oduct of a disease resistance
gene (R) in th~ plant and the product of a
corresponding avirulence (Avr) gene in the
pathogen. A plant variety can be transformed
with cloned resistance gene to engineer plants
that are resistant to specific pathogen
strains. See, fog example Jones et al.,
Science 265:789 (1994) (cloning of the tomato
Cf-9 gene fo= resistance to Cladosporium
fulvum) ; Mar ti n et a1 . , Science 262 :1432 (1993)
(tomato Pto aen~ ~o= resistance to Pseudomonas
syringa= pv. tomato encodes a protein kinase);
Mindrinos et a?., Cel.I 78:1089 (1994)
CA 02355198 2001-07-18
X29-137
17
(Arabidopsis RSP2 gene for resistance to Pseudomonas
syringa2) .
(B) A Bacillus thuringiensis protein, a derivative
thereof or a synthetic polypeptide modeled thereon. See, for
example, Geiser et al., Gene 48:109 (1986), who disclose the
cloning and nucleotide sequence of a Bt 8-endotoxin gene.
Moreover, DNA molecules encoding b-endotoxin genes can be
purchased from American Type Culture Collection (Rockville,
MD), under ATCC accession Nos. 40098, 67136, 31995 and 31998.
(C) A lectin. See, for example, the disclosure by
Van Damme et al., Plant Molec. Biol. 24:825 (1994), who
di,~close the nucleotide sequences of several Clivia miniata
mannose-binding lectin genes.
(D) A vitamin-binding protein such as avidin. See
PCT patent publication serial No. W09400992. The application
teaches the use of avidin and avidin homologues as larvicides
against insect pests.
An enzyme inhibitor, for example, a protease
inhibitor or an amylase inhibitor. See, for example, Abe
et al., J. Biol. Chem. 262:16793 (1987) (nucleotide sequence of
rice cysteine proteinase inhibitor), Huub et al., Plant Molec.
Biol. 21:985 (1993) (nucleotide sequence of cDNA encoding
tobacco proteinase inhibitor I), and Sumitani et al., Biosci.
Biotech. Biochsm. 57:1243 (1993) (nucleotide sequence of
Strsptomyces nitrosporsus cc-amylasa inhibitor).
(F) An insect-specific hormone or pheromone such as
an ecdysteroid and juvenile hormone, a variant thereof, a
mimetic based thereon, or an antagonist or agonist thereof.
See, for
CA 02355198 2001-07-18
96!30530 PCTIUS96I03.
_ i8 _
example, the disclosure by Hammock et al.,
Nature 344:458 (1990), of baculovirus
expression of cloned juvenile hormone esterase,
an inactivator of juvenile hormone.
S (G) An insect-specific peptide or neuropeptide
. which, upon expression, disrupts the physiology
of the affected pest. For example, see the
disclosures of Regan, J. Biol. Chem. 269:9
(1994) (expression cloning yields DNA coding
for insect diuretic hormone receptor), and
Pratt et al., Biochem. Biophys. Res. Comm.
163:1243 (1989) (an allostatin is identified in
Diploptera puntata). See also U.S. patent No.
' 5,266,317 to Tomalski et al., who disclose
genes encoding insect-specific, paralytic
neurotoxins.
(H) An insect-specific venom produced in nature by
a snake, a wasp, etc. For example, see Pang et
al., Gene 116:i6S (1992), for disclosure of
heterologous e:<pression in plants of a gene
coding for a scorpion insectotoxic peptide.
(I) An enzyme responsible for an hyperaccumulation
of a monterpen~, a sesquicerpene, a steroid,
hydroxamic acid, a phenylpropanoid derivative
or another non-protein molecule with
insecticidal activity.
(J) An enzyme involved in the modification,
includin3 the pose-~~anslational modification,
of a biologically ac~ive molecule; for example,
a glycolytic enzyme, a proteolytic enzyme, a
lipolytic enzyme, a nuclease, a cyclase, a
transaminase, an esterase, a hydrolase, a
phosphacase, a kinase, a phosphorylase, a
polymerase, an elascase, a chitinase and a
glucanase, whech~r natural or synthetic. See
PCT applicatio:: ~rlC 73/02197 in the name of
CA 02355198 2001-07-18
29-137
19
Scott et al., which discloses the nucleotide sequence of a
callase gene. DNA molecules which contain chitinase-encoding
sequences can be obtained, for example, from the ATCC under
accession Nos. 39637 and 67152. See also Kramer et al., Insect
Biochem. Molec. Biol. 23:691 (1993), who teach the nucleotide
sequence of a cDNA encoding tobacco hookworm chitinase, and
Kawalleck et al., Plant Molec. Biol. 21:673 (1993), who provide
the nucleotide sequence of tie parsley vbi4-2 polyubiquitin
gene.
(K) A molecule that stimulates signal transduction.
For example, see the disclosure by Botella et al., Plant Mole.
Biol. 24:757 (1994), of nucleotide sequences for mung bean
c~:lmodulin cDNR clones, and Griess et al., Plant Physiol.
104:1467 (1994), who provide the nucleotide sequence of a maze
calmodulin cDNA clone.
(L) A hydrophobic moment peptide. See U.S. patent
serial No. 5,580,852 (disclosure of peptide derivatives of
Tachyplesin which inhibit fungal plant pathogens) and serial
No. 5,607,914 (teaches synthetic antimicrobial peptides that
confer disease resistance).
(M) A membrane permease, a channel former or a
channel blocker. For example, see the disclosure by Jaynes
et al., Plant Sci. 89:43 (1993), of heterologous expression c.
a cecropin-~ lytic peptide analog to render transgenic tobac~~
plants resistant to Pseudomonas solanacearvm.
(N) A viral-invasive protein or a complex toxin
derived therefrom. For example, the
CA 02355198 2001-07-18
.~ 96/30530 PCTIUS96/0~.._..
- 2C -
accumulation of viral coat proteins in
transformed plant cells imparts resistance to
viral infection and/or disease development
effected by the virus from which the coat
protein gene is derived, as well as by related
viruses. See Beachy et al., Ann. Rev.
Phytopathol._ 28:451 (1990). Coat protein-
mediated resistance has been conferred upon
transformed plants against alfalfa mosaic
virus, cucumber mosaic virus, tobacco streak
virus, potato virus X, potato virus Y, tobacco
etch virus, tobacco rattle virus and tobacco
mosaic virus. Id.
(O) An insect-specific antibody or an immunotoxin
derived therefrom. Thus, an antibody targeted
to a critical metabolic function in the insect
gut would inactivate an affected enzyme,
killing the insect. Cr. Taylor et al.,
Abstract r497, S~V~NTH INT'L SYMPOSIUM ON
MOLECULA.? PLAiVT-f'ICROB~ INTERP,CTIONS (1994)
(enzymatic inactivation in transgenic tobacco
via production o' single-chain antibody
fragments) .
(P) A virus-specific a:~tibody. See, for example,
Tavlado=aki et al., Natur= 366:469 (1993), who
show that transg=nit plants expressing
recombinant antibody genes are protected from
virus attack.
(Q) A developmental-arrestive protein produced in
nature by a pathogen or a parasite. Thus,
fungal endo cr-1,4-D-polygalacturonases
facilitate funga'_ colonization and plant
nutrient release by solubilizing plant cell
wall homo-a-1,4-D-galacturonase. See Lamb et
al., 9io/Technoloay 10:1436 (1992). The
cloning and characterization of a gene which
CA 02355198 2001-07-18
96!30530 PCTlUS96103.
-, .
_ c. _
encodes a bean endopolygalac-turonase-
inhibiting protein is described by Toubart et
al., Plant J. 2:367 (1992).
(R) A developmental-arrestive protein produced in
nature by a plant. For example, Logemann et
al., Bio/Technology 10:305 (1992), have shown
that transgenic plants expressing the barley
ribosome-inactivating gene have an increased
resistance to fungal disease.
2. Genes That Confer Resistance To A Herbicide For
Example:
(A) A herbicide that inhibits the growing point or
~ . meristem, such as an imidazalinone or a
sulfonylurea. Exemplary genes in this category
code for mutant ALS and AHAS enzyme as
described, for examqle, by~Lee et al., EMBO J.
7 : 1241 ( 1988 ) , ar.~d ~Iiki et a1 . , Theor. Appl .
Gene . 80:449 (1990?, respectively.
(B) Glyphosate (resistance imparted by mutant EPSP
synthase and aro.4 genes, respectively) and
other phosphono co.a,pounds such as glufosinate
(PAT and bay genes;, and pyridinoxy or- phenoxy
proprionic acids an:: cycloshexones (ACCase
inhibitor-encodinc genes). See, for example,
U.S. patent No. 4,340,835 to Shah et al., which
discloses the nucleotide sequence of a form o.
EPSP which can core= glyphosate resistance.
A DNA molecule encoding a mutant aroA gene can
be obtained under ATC'C accession No. 39256, and
the nucleotide secu_nce of the mutant gene is
disclosed in U.S. patent No. 4,769,061 to
Comai. European gatent application No. 0 333
033 to Kumada ~~ a~. and U.S. patent No.
4, 975, 37-~.~. to Goodman et a1. disclose nucleotide
sequences oL alLtam=ne synthetase genes which
CA 02355198 2001-07-18
96130530 PCTIUS96I03w
- 2~ -
confer resistance to herbicides such as L-
phosphinothricin. The nucleotide sequence of
a phosphinothricin-acetyl-transferase gene is
provided in European application No. 0 242 246
to Leemans et a1. ~De Greef et al.,
Bio/Tecnnology 7:61 (1989), describe the
production of transgenic plants that express
chimeric bar~genes coding for phosphinothricin
acetyl transferase activity. Exemplary of
genes conferring resistance to phenoxy
proprionic acids and cycloshexones, such as
sethoxydim and haloxy=op, are the Acc1-S1,
Acct-S2 and Accl-~3 genes described by Marshall
' et al., Theor. ADD1. Genet. 83:435 (1992).
(C) A herbicide that i::hibits photosynthesis, such
as a triazine (psbA and gs+ genes) and a
benzonitrile (nit:ilase gene). Przibilla et
al., Planc Cell 3:10'9 (1991), describe the
transformation o~ !'::lamydomonas with plasmids
encodirc mutant psb.4 genes. Nucleotide
sequences for ~=t_~?ase genes are disclosed in
U.S. patent No. 4,310,548 to Stalker, and DNA
molecules containinc these genes are available
under ATCC acc~ssio:~ Nos. 53435, 67441 and
67442. Cloning and expression of DNA coding
for a giu.tathione ~-transferase is described by
Hayes et al., 9io'nem. J. 285:173 (1992).
3. Genes That Confer Or Contribute To A Value-Added
Trait, Such As:
(A) Modified Latty ac_;~etabolism, for example, by
transforming maiz=_ 3rassica with an antisense
gene of stearoyl-~C? desaturase to increase
stearic aci~ co.~.t~:.t o. the plant . See
Knultzoz ec a1. , P=cue. lVat' 1 Acad Sci. USA
89:2624 (19921.
CA 02355198 2001-07-18
~ 96130530 PCT/US96/0~.
- 23 -
(B) Decreased phytate content
(1) Introduction of a phytase-encoding gene
would enhance breakdown of phytate, adding
more free phosphate to the transformed
plant. For examDle,~see Van Hartingsveldt
et al., Gene 127:87 (1993), for a
disclosure of the nucleotide sequence of
an Aspergillus niger phytase gene.
(2) A gene could be introduced that reduces
phytate content. In maize, this, for
example, could be accomplished, by cloning
and then ~_-introducing DNA associated
with the singly allele which is
responsible for maize mutants
characterized by low levels of phytic
acid. See ~aboy et al., Maydica 35:383
(1990) .
(C) Modif'_ed carbohydrate composition effected, for
example, by transforming plants with a gene
coding fo= a~ e::zym~ that alters the branching
patte_n of start~. See Shiroza et al., J.
Bact~riol. 170:810 (1988) (nucleotide sequence
of Streptococcus mutans fructosyltransferase
gene), Steinmetz et al., Mol. Gen. Genet.
200:220 (1985) (nucleotide sequence of Bacillus
subtilis levansucrase gene), Pen et al.,
Bio/Tecnnology 10:292 (1992) (production of
transgenic plants shat express Bacillus
lichenirormis or-amylase), Elliot et al., Plar.t
Molec. Biol. 21:S1S (1993) (nucleotide
sequences o= tomato invertase genes), SOgaard
et a:.., J. Biol. Chum.. 268:22480 (1993) (site-
directed mutagenesis of barley amylase gene),
and :ishe=- et a'., Plant Physiol. 102:1045
(1993) (ma_ze endosperm starch branching enzyme
II) .
CA 02355198 2001-07-18
.. ~! 96130530 PCT/US961030.,~
- 2~-_ -
4. Selectable Marker Genes:
(A) Numerous selectable marker genes are available
for use in plant transformation including, but
not limited to, neomycin~.phophotransferase I,
S hygromycin phophotransferase, EPSP synthase and
dihydropteroate. See Miki et al., "Procedures
for Introducing Foreign DNA into Plants," in
METHODS IN PLANT MOLECULAR BIOLOGY AND
BIOTECHNOLOGY, Glick et aI. (eds.), pages 67-88
(CRC P2-ess, 1993) .
Synthesis of g=nes suitably employed in the present
invention can be e_~ect~d by mans or mutually priming,
long oligonucleotides. Se" Lor example, Ausubel et a1.
(eds.), CURRENT PROTOCOLS IN htOLECUI~AR BIOLOGY, pages
8.2.8 to 8.2.13 (Wiley Inte=science 1990) , and Wosnick et
al., Gene 60:115 (1987). Mo_eover, current techniques
which employ the polvmerase chain reaction permit the
synthesis or genus as large as 1.8 kilobases in length.
See Adang et al., Plant hlol°c. 9iol. 21:1131 (1993), and
Bambot et a1. , PCR :~hthods and ~.pplications 2:266 (1993) .
The present invention, thus generally described, will
be understood more =eadily by =eference to the following
examples, which are provided by way of i'_lustration and
are not intended to be limiting of the present invention.
Example I
Plant Trans~ormation and expression Analysis
The vector p?::I4960, shown in Figure 3, was
introduced into the Agrobacterium strain GV3101 by
transformation. S. rapes cultivar Westar was used
throughout this experiment. Transgenic B. napes plants
were generated by Agro.bacterium co-cultivation of
cotyledonary petioles and microspore derived embryos.
See Arnoldo et al., G~ne:n~ 35:58 (1992). Parallel
transformations w~r~ carr~~d out using pPHI3042 in order
to compare the Stre:lgt:1 G. ~h~ ALS3 prompter with that of
CA 02355198 2001-07-18
~ 96/30530 PCT1US96/0~_
_ 25 _
the CaMV 35S promoter. Plasmid pPHI3042 is shown in
Figure S and contains the CaMV 35S promoter operably
linked to the GUS gene. Tissues from plants containing
independent transformation events were quantitatively
analyzed for GUS expression using a histochemical assay
method based on-that of Jefferson R.A., Plant Mol. Biol.
Rep. 5:387 (1987).
Table 1 summarizes quantitative GUS expression data
from B. napes plants independently transformed with
_ plasmid pPHI4960 (HWSC63E, HWSC63F, HWSC63G, HWSC63H,
PWSC174C, PWSC174D, PWSC174G, PWSC174J, PWSC174M,
PWSC174N, and PWSC176A). Tissue from the primary
transgenic plant (TO) was analyzed. Plasmid pPHI4960 is
shown in Figure 3 and contains the 9. napes ALS3 promoter
operably linked to the GUS acne. The controls consisted
of the same nontransformed cultivar 'Westar' and 4
independent B. napes plants __ansformed with the plasmid
pPHI3042 (P10SB, P105=, P1253 and P145).
CA 02355198 2001-07-18
~6I30530 PCT/US96/03a.
_ 2~ _
Table 1
SUMMARY OF BRASSICA NAPUS TRANSFORMED
WITH pPHI4960 AND pPH13042
Transgenic GUS Expression.(TO) Southern
Fluorogenic PI1VII Probe
(pmoles/MU/h/~cg j
protein) '
leaf root
HWSC63E~ 0.2 6.2 5
HWSC63F~ 131 . 167 1
HWSC63G~ 416. ( 246 1
HWSC63H~ 21 29 1
PWSC174C~ 7.S 63.6 1
PWSC174D~ 11 I 17 1
PWSC174G~ 309 nc root 0/1
PWSC174J~ 1
PWSC174M~ 0/1
PWSC174N~ 1
PWSC176A~ 0.2 37 4
P105B' 315 I 143
P10SEZ 24. f 279
P126B2 0 . -~. ~ 1 . 2
Pl4Sz 0.2 I 0.6
WESTAR3 0.1 I 11.2
B. napes trans~orm~d wig:z pPH_T4960
B. napes trans:.ormed wi~:z oPH:3042
Non-transgenic co::crc
CA 02355198 2001-07-18
96130530 PCTIUS9510_
- 27 -
The data reveal that there is variability in the
level of expression in the transformants. Variability in
the level of expression of transformed genes has been
noted in numerous transformation experiments described in
the literature and has been attributed to such factors as
differences in copy number, position effects and
co-suppression. However, plants expressing high levels,
or a desired level, of the foreign protein can' be
identified and selected using the routine screening
methods of the present invention.
The data also reveal that there are essentially two
classes of expression levels in plants transformed with
plasmid pPHI4960. The first class consists of
transformed plants expressing high levels of assayable
product (131-416 pmal of methylumbelliferone (MU)/h/~eg
protein). The second class consists of transformed
plants expressing 1 ow levels of assayable product (0 .2-37
pmol MU/h/~g protzin). The low expression level is
generally comparable to the non-transformed control
plants examined. These values do not imply absolute
expected ranges but represent the maximum and minimum
values for each class in the present study.
Southern blot analysis of total genomic DNA isolated
from transformed ?ants was undertaken to determine the
copy number of the GUS gene cassette in each
transformant. The PINI_ terminator was used as the
radiolabeled hybridization probe. Total genomic DNA was
isolated from eacr plant transformant using a variation
of a CTAB protocol. See, .or example, Dellaporta et al.,
1983, Plant MoI. Biol. Reo. 1(4):19-21, or Saghai-Maroot
et al., 1984, PNAS 8:8014-8018. The total genomic DNA
was digested with a restriction enzyme(s), separated on
agarose gels by means of electrophoresis and hybridized
with the radiolab_led PINIT terminator sequence as a
~ probe. Plasmid pP:iI4960 was digested with either HindIII
alone to assess number o= irt_grations, or double
digested with Hindi=T plus EcoRI to determine copy
number. Similarly, pP:-iI3042 was digested with HindIII
CA 02355198 2001-07-18
X6130530 PCTIUS96/03.
- 28 -
alone to assess number of integrations, or with EcoRI
alone to evaluate copy number (see enzyme cut sites on
maps in Figures 3 and 5, respectively). Although an
exact copy number for each integration event was not
determined, the relative intensities of hybridizing bands
were used to compare Southern data between transgenics
with different vectors.
Plants transformed with multiple copies of the
plasmid pPHI4960 (HWSC63E~and PWSC176A) produced low
levels of MU. On the other hand, 50% of the single
integration events produced high levels of MU. Plants
with single transformation events produced high levels of
MU irrespective of whether the GUS gene was expressed
from the CaMV 35S or the ALS3 promoter.
15~ ' Among the transformed plants producing high levels
of MU, plants transformed with the ALS3 GUS cassette
produced just as much MU as plants transformed with the
CaMV 35S GUS cassette, irrespective of whether enzyme
activity was measured in the roots or shoots. Since the
CaMV 35S promoter is regarded as a strong constitutive,
generally non-tissue-preferred promoter, with wide
application in expressing genes in transgenic plants, the
Brassica ALS3 promoter offers a suitable alternative to
the CaMV 35S promoter.
The level of GUS expression measured in leaf tissue
of HwSC63G was higher than the GUS level produced in leaf
or root tissue of plants transformed with the CaMV 35S
GUS cassette. Accordingly, the ALS3 promoter may be mor=_
effective than the CaMV 35S promoter for expression of
certain genes at high levels. For example, the ALS3
promoter may be preferable to the CaMV 35S promoter in
those instances where high level expression of the
transformed gene is required. It is understood by those
skilled in the art that the range of expression in
transgenic plants can vary among transformation events.
The present data indicates that it is possible to recover
at least some events with activity comparable to that
CA 02355198 2001-07-18
. .. .~ 96130530 PCT/US96/0~_ _
- 29 -
obtained with the 35S promoter by means of routine
screening using the methods disclosed herein.
To test the usefulness of the ALS3 promoter for
expression of operably linked genes in other species,
transformations were also conducted in sunflower.
Sunflower leaf discs were ~~co-cultivated with
Agrobacterium carrying either pPHI4960 or pPHI6333. See
Malone-Schoneberg, et al., 1994, Plant Science 103: 199-
207. Stably transformed calli were selected using 100
mg/L of kanamycin. GUS or NPTII expression levels were
determined for a number of independent transformation
events, using histochemical staining for GUS expression
analysis, and semi-quantitative ELISAs for NPTII
. expression analysis. While GUS expression varied among
individual plants, the ranges of expression from the two
promoters in leaf tissues overlapped substantially (73
3119 fluorescence units for the ALS3 promoter and 40-2788
fluorescence units for the 35S promoter) . Limited ELISAs
also yielded comparable results for NPTII expression from
the two promoters (data not shown), although extensive
whole plant analysis was not carried out.
Example 2
Activity of the ALS3 Promoter in Transgenic 9. napus ir.
Selected Tissues from Seedling, vegetative, Flowering
and Mature Stage Plants
B. napus cv. Westar was transformed with DPHI4960 as
described in Example 1. Four independent transformants
were analyzed (HWSC63F, HWSC63G, HWSC63E and PWSC176A).
The primary transformants are designated T0. The seed
produced by TO plants are T1 seeds and the plants
produced by T1 seeds are referred to as T1 plants.
GUS expression was quantitatively determined, by the
method described in Example 1, in T1 plants at t
seedling stage (less than 2 true leaves), the vegetative
stage (4-5 true leaves) , the early flowering stage (about
20-80% full bloom), and in mature plants. In addition,
GUS expression was quantitatively determined in specific
plant tissues, including meristem, cotyledon, hypocotyl,
CA 02355198 2001-07-18
96130530 PCT/US96J0~.
- 30 -
leaf, petiole, stem, root, pod, seed and flowers.
"Meristem" refers to the meristematic apex and includes
much of the tissue surrounding the true meristem.
"Flowers" refers to all organs associated with the
flowers, including ovaries, anthers and petals. "Pods"
refers to the developing green pods containing green and
brown seed. "Seed" refers to the seed still on the
plant, hence the seeds were not desiccated. The results
of quantitative enzyme assays were confirmed by means of
histochemical assays as in example 1.
GUS expression in B. napes cv. Westar transformed
with pHI4960 was compared to plants of the same cultivar
transformed with pPHI3042. A non-transgenic Westar line
was used as a control. Although T1 stage plants are
segregating with respect to the transformed genes, only
T1 plants that had a transgene, as evidenced by
expression of neomycin phosphotransferase or survival on
kanamycin, were analyzed. Seeds were sown on an agar
plate containing 100 ~g/ml of kanamycin. Those seeds
which germinated and resulted in green seedlings were
selected as transformed segregants. Those which bleached
white were non-transformed segregants and were discarded
or used as negative controls.
Table 2 summarizes Southern, expression and
segregation data for several transgenics. GUS expression
was measured in leaf tissue from plants at the 3-7 leaf
stage. In those primary transgenics with a simile
integration pattern, which either showed no segregation
(and were therefore fixed) o. whose subsequent progeny
segregated in approximately a 3:1 ratio with respect to
the transformed genes, the levels of GUS expression from
both the 35S and ALS3 promoters was relatively high. In
contrast, those with complex integration patterns and/or
with complex segregation had low GUS expression.
CA 02355198 2001-07-18
X6130530 PCTIUS96/03.
- 31 -
a O
o ~o w c ; i
aQ ...00 ? ~ ~ O
e'1i LL ... v
.
v
v ~ _ v
N _ 'r _
N t~
o ~ x
.
.
0 ~ ~ o
o .
x -.~ ~ z
U
E
.d
N
JJ
O
x ~ ~ ~ x
3 3
~
w a ~
x _ x
c~
0
C:1~ U w w w ~ L7
s z Z Z Z z z Z z
o _' _ _ -~ _.
a N v v~
~
a a ~ ~ a
a e -,
U ,
'n
U ~ U E~ U U U U
3 ~ 3 ~ ~ 3 3 3 3
z z ~ < o. x _ s
CA 02355198 2001-07-18
16130530 PCTlUS96I03~
- 32 -
Figures 7, 8, 9 and 10 present GUS expression data
in specific tissues taken from plants at the seedling,
vegetative, flowering and mature stage, respectively. As
evidenced by the results shown in Figure 7, the ALS3
S promoter functions in seedlings, specifically in
cotyledons, hypocotyls and "meristeinatic' apex. " The ALS3
promoter drives low level expression of GUS in the roots
at the seedling stage. At the 4-5 true leaf stage,~the
ALS promoter functions in leaves, petioles, stem,
"meristematic apex" and roots, as shown in Figure 8. At
the flowering stage, the ALS3 promoter functions in
flowers, leaves, stem and roots (Figure 9).
Histochemical analysis of flowers revealed expression of
GUS in petals, stigma, anthers and pollen. GUS activity
1S was detected in all tissues (leaf, stem pod and seed)'
taken from mature B. napes plants transformed with
pPHI4960. Accordingly, the ALS3 promoter is
constitutively expressed in diverse plant tissues
throughout the development of the plant.
Exam~Ie 3
Selectable Marker Genes Expressed
from the ALS3 Promoter
The ALS3 promoter can be used to dr ive expression of
selectable marker genes for plant transformation. B.
2S napes was transformed with plasmids pPHI1741, pPHI3042,
pPHI576S and pPHI6333 (Figure 4) using the methods
described in Example 1. Transformants were selected on
medium containing 100 ~.g/ml kanamycin. Plasmid pPHI1741
contains the CaMV 3SS promoter operably linked to the
NPTII structural gene and the terminator sequence of the
19S gene. Plasmid pPHI3042 contains two expression
cassettes with the CaMV 3SS promoter operably linked to
the NPTII structural gene and the CaMV 3SS promoter
operably linked to the GUS gene. Plasmid pPHI576S
3S contains the CaMV 3SS promoter operably linked to the
NPT'II structural gene. Finally, plasmid pHI6333, shown
in Figure 4, contains the ALS3 promoter operably linked
CA 02355198 2001-07-18
.. ~ 96/30530 PCTlUS96f03~_.,
- 33 -
to the NPTII structural gene and the terminator sequence
of the potato proteinase inhibitor gene (PINII).
Table 3
Selectable Marker Genes Expressed
From the ALS3 Promoter
TOTAL NUMBER TOTAL PERCENT
VECTOR OF EXPLANTS NUMBER OF TRANS-
COCULTIVATED POSITIVES FORMED
pPHI1741 1350 64 4.7%
' pPHI3042 350 2 0.9%
pPHI5765 1935 60 3.1%
pPHI6333 1935 12 0.6%
As shown in Table 3, the ALS3 promoter is effective
in driving expression of the NPT'II selective marker gene
in canola. Transformation of plasmids pPHI1741, pPH3042
and pPHI5765 resulted in the recovery of 4.7%, 0.9% and
3.1% transformants among the total B. napus explants
cocultivated with these plasmids, respectively. A total
of 12 transformants were obtained from experiments in
which pPHI6333 was cocultivated with 1,935 B. napus
explants, or 0.6% of the explants were transformed. Ten
of the 12 transformants expressed low levels of NPTII.
In sunflower, pPHI6333 was as effective as pPHI4960 for
selection of transformed plants; that is, both the 35S
and the ALS3 promoter allowed selection of transformed
tissues at 100 mg/L of kanamycin. Accordingly, the ALS3
promoter is effective in driving expression of a
selective marker gene for plant transformation.
Although the foregoing refers to particular
preferred embodiments, it will be understood that the
CA 02355198 2001-07-18
'9-137
34
present invention is not so limited. It will occur to those of
ordinary skill in the art that various modifications may be
made to the disclosed embodiments and that such modifications
are intended to be within the scope of the present invention,
which is defined by the following claims.
All publications and patent applications mentioned in
this specification are indicative of the level of skill of
those in the art to which the invention pertains.