Note: Descriptions are shown in the official language in which they were submitted.
CA 02507563 2005-05-13
Express 1V' Label No. EV2895101 ~SUS
PATENT APPLICA'f 10N
ATTORNEY DOCKET NO.: GOW-001 CP2
PLANT REGULATORY ELEMENT
[0001] This application is a continuation-in-part of U.S.S.N. 09/457,123,
filed
December 7, 1999, which is a continuation-in-part of U.S.S.N. 09/174,999,
filed
October 19, 1998, now abandoned, which is a continuation of U.S.S.N.
08/593,I2I,
filed February 1, 1996, now U.S. Patent No. 5,824,872, issued October 20,
1998, the
entire disclosures of each of which are incorporated herein by reference.
FIELD OF INVENTION
[0002] The present invention relates to regulatory elements obtained from a
plant.
This invention further relates to the use of one or more than one regulatory
eler.ent to
control the expression of exogenous DNAs of interest in a desired host.
BACKGROUND OF THE INVENTION
(0003] Bacteria from the genus Agrobacterium have the ability to transfer
specific
segments of DNA (T-DNA) to plant cells; where they stably irte~~ ate i;,to t
he r~aclear
chromosomes. Analyses of plants harbouring the T-DNA have revealed that this
genetic element may be integrated at numerous locations, and can occasionally
be
found within genes: One strategy which has been exploited to identify
integration
events within genes is to transform plant cells with specially designed T-DNA
vectors
which contain a reporter gene, devoid of cis-acting transcriptional and
translational
expression signals (i.e. promoterless), located at the end of the T-DNA. Upon
integration, the initiation codon of the promoterless gene (reporter gene)
will be
juxtaposed to plant sequences. The consequence of T-DNA insertion adjacent to,
and
downstream of, gene promoter elements may be the activation of reporter gene
expression. The resulting hybrid genes, referred to as T-DNA-mediated gene
fusions,
consist of unknown and thus un-characterized plant promoters residing at their
natural
location within the chromosome, and the coding sequence of a marker gene
located on
the inserted T-DNA. (Fobert et al., 1991, Plant Mol. Biol. 17, 837-851 ).
CA 02507563 2005-05-13
[0004] It has generally been assumed that activation of promoterless or
enhancerless
marker genes result from T-DNA insertions within or immediately adjacent to
genes.
The recent isolation of several T-DNA insertional mutants (Koncz et al., 1992,
Plant
Mol. Biol. 20, 963-976; reviewed in Feldmann, 1991, Plartt J. 1, 71-82; Van
Lijsebettens et al., 1991, Plant Sci. 80, 27-37; Walden et al., 19915 Plant J.
1:
281-288; Yanofsky et al., 1990, Nature 346, 35-39), shows that this is the
case for at
least some insertions. However, other possibilities exist. One of these
possibilities is
that integration of the T-DNA activates silent regulatory sequences that are
not
associated with genes. Lindsey et al. (1993, Transgenic Res. 2, 33-47)
referred to
n ,a~ +~ v ,~ F,.,7 aL..a aL..__ t-,. o_t r_..
,iW a ~e~iW.ui.v.$ aJ YWuuv-yrvuivwrS aiiu Su~gcS~cu maW ,itcy 111~t)' UG
leJ~JU11J1U1e. lUI
activating marker genes in some transgenic lines. Fobert et al. (1994, Plant
J. 6, 567-
577) have cloned such sequences and have referred to these as "cryptic
promoters".
[0005] Mandel et al (1995, Plant Molec. Biol. 29:995-1004) discloses a
promoter
which is active in leaves, stem, and apical meristem tissues. This promoter
was
obtained from translation initiation factor 4A (NeIF-4A), a house keeping gene
found
+.,1.,.,1 11. > +:..0 11
in Wemuvrl arty aCmv. CvllJ.
[0006] Other regulatory elements are located within the 5' and 3' untranslated
regions
(UTR) of genes. These regulatory elements can modulate gene expression in
plants
through a number of mechanisms including translation, transcription and RNA
stability. For example, some regulatory elements are known to enhance the
translational efficiency of mRNA, resulting in an increased accumulation of
recombinant protein by many folds. Some of those regulatory elements contain
translational enhancer sequences or structures, such as the Omega sequence of
the 5'
leader of the tobacco mosaic virus (Gallie and Walbot, 1992, Nucleic Acid res.
20,
4631-4638), the 5' alpha-beta leader of the potato virus X (Tomashevskaya et
al, 1993,
J. Gen. Virol. 74, 2717-2724), and the 5' leader of the photosystem I gene
psaDb of
Nicotiana sylvestris (Yamamoto et al., 1995, J. Biol. Chem 270, 12466-12470).
Other
5' regulatory elements affect gene expression by quantitative enhancement of
transcription, as with the UTR of the thylakoid protein genes PsaF, PetH and
PetE
2
CA 02507563 2005-05-13
from pea (Bone et al., 199, Plant J. 6, 513-523), or by repression of
transcription, as
for the S' UTR of the pollen-specific LATS9 gene from tomato (Curie and
McCormick, 1997, Plant Cell 9, 2025-2036). Some 3' regulatory regions contain
sequences that act as mRNA instability determinants, such as the DST element
in the
Small Auxin-Up RNA (SAUR) genes of soybean and Arabidopisis (Newman et al., .
'
1993, Plant Cell S, 701-714). Other translational enhancers are also well
documented
in the literature (e.g. Helliwell and Gray 1995, Plant Mol. Bio. vol 29, pp.
621-626;
Dickey L.F. al. 1998, Plant Cell vol 10, 475-484; Dunker B.P. et al. 1997 Mol.
Gen. ~ '
Gent. vol 254, pp. 291-296).
SUi~Ii~IARY OF THE INVENTION
[0007) The present invention relates to regulatory elements obtained from a
plant.
This invention further relates to the use of one or more than one regulatory
element to
control the expression of exogenous DNAs of inteiest in a desired host.
[0008] It is an obect of the irwention to provide an improved constitutive
regulatory
element.
[0009] The transgenic tobacco plant, T127S, contained a 4.38 kb EcoRIlXbaI
fragment containing the 2.1 S kb promoterless GUS-nos gene and 2.23 kb of 5'
flanking tobacco DNA (2225 bp). This 5' flanking DNA shows no homology to
known sequences, and exhibits constitutive regulatory element activity.
Analysis of
the S' flanking DNA revealed the occurrence of several additional regulatory
elements,
and that this DNA is a member of a large family of repetitive elements.
[0010] The present invention relates in part to an isolated plant constitutive
regulatory
element that directs expression in at least ovary, flower, immature embryo,
mature
embryo, seed, stem, leaf, root and cultured tissues of a plant. preferably,
the
regulatory element is not obtained from a IFA-4A gene. The isolated plant
-,
J
CA 02507563 2005-05-13
constitutive regulatory element may also be characterised by lacking an intron
in its
5'UTR and a TATA box.
[0011 ] The constitutive regulatory element could not be detected in soybean,
potato,
sunflower, Arabidopsis, B. napus, B. oleracea, corn, wheat or black spruce by
Southern blot analysis. However, expression of a coding region of interest,
under
control of the regulatory element, or a fragment thereof, was observed in
transgenic
tobacco, N. tabacum c.v. Petit Havana, SRI, transgenic B. napus c.v. Westar,
transgenic alfalfa, and transgenic Arabidopsis, and Was observed in leaf,
stem, root,
developing seed and flower. In transient expression analysis, GUS activity was
also '
observed ir~ leaf tissue of soybean, alfalfa, Arabidopsis, tobacco, B. napus,
pea, potato,
peach, Ginseng and suspension cultured cells of white spruce, oat,
cern,~~Nheat and
barley.
[0012] Thus this invention also provides for a regulatory element that is a
constitutive
regulatory element. Furthermore, this regulatory element functions in diverse
plant
species when introduced on a cloning vector, and maybe used to drive the
expression
of a coding region of interest within a range of plant species.
[0013] The present invention also relates to an isolated plant regulatory
element that
directs expression in at least ovary, flower, immature embryo, mature embryo,
seed,
stem, leaf, root and cultured tissues of a plant, wherein the regulatory
element, or a
fragment thereof, is a repetitive element. Preferably, the isolated plant
regulatory
element is a member of the RENT family of repetitive elements.
[0014] This invention pertains to a regulatory element characterized in that
it
comprises at least an 18 by contiguous sequence of any one of SEQ ID NO's:l,
5, 6,
7, 8, 9, 21 and 22.
[0015] The present invention also embraces a regulatory element having a
nucleotide
sequence that hybridizes to a nucleotide sequence, or a fragment thereof, as
defined by
the nucleotide sequence of any one of SEQ ID NO: 1, ~, 6, 7, 8, 9, 21 and 22
under the
4
CA 02507563 2005-05-13
following hybridization conditions: 4XSSC at 65°C overnight, followed
by washing
in O.1XSSC at 65°C for one hour, or twice for 30 minutes each, wherin
the nucleotide
sequence exhibits regulatory element activity.
[0016] The transcription start site for the introduced GUS gene in transgenic
tobacco
was located in the plant DNA upstream of the insertion site. It was the same
in leaf, ~ ~ , '
stem, root, seeds and flower. Furthermore; the .native site was silent in both
, '
untransformed and transgenic tobacco. ' ' , ~ ' ' ' ,
[0019] This invention also relates to a chimeric construct comprising a coding
region .
F:.,+Q..o + F -. .1.:,.1. +:+_,+:~.~ - a.-.-._--a a - ~ '
W mwW~ m ~vmvu witJW uuW . 2Ailli~JJ1V11, 1J lLGJ11G11, cL11C1 Q
I~UIl~LllltllV~ ~C~LIIaLUTy
element, comprising at least an 18 by contiguous sequenYe of ar~y one SEQ ID
NO's:
1, 5; 6, 7, 8, 9, 2i and 22. This invention further relates to a cloning
vector containing
the chimeric gene construct. ' ,
[0018] This invention also includes a plant cell which has been transformed
with the
chimeric gene, or cloning vector as defined above: Furthermore, this invention
a ),rarac tranogaroir~ p1."-,t$~ ~rld seeds, voiltaiiling t~le 1. hi111Gr1C
geile, Or tile 1:1V111I1g
ri'..u. uvv.. u.p ...aw. iuia
vector as defined above.
[0019] This invention further relates to any transgenic host, for example, but
not
limited to a transgenic plant, containing a nucleotide sequence selected from
the group
consisting of SEQ ID NO's: 1, 5, 6, 7, 8, 9, 21 and 22 or nucleic acid
sequence that
hybridizes to the nucleotide sequence, a complement, or a fragment thereof, as
defined
by the nucleotide sequence of any one of SEQ ID NO's: 1, 5, 6, 7, 8, 9, 21 and
22
under the following hybridization conditions: 4XSSC at 65°C overnignt,
followed by
washing in O.1XSSC at 65°C for one hour, or twice for 30 minutes each.
The
nucleotide sequence may also be operatively linked to a coding region of
interest that
is transcribed into RNA. Preferably, the coding region is heterologous with
respect to
the regulatory region.
CA 02507563 2005-05-13
[0020] Also included in the present invention is a method of conferring
expression of
a coding region of interest in a plant, comprising: operatively linking an
exogenous
coding region of interest, for which constitutive expression is desired, with
a
regulatory element comprising at least an 18 by contiguous sequence of any one
of
SEQ ID NO's:l, 5, 6, 7, 8, 9, 21 and 22 to produce a chimeric construct and
introducing the chimeric construct into a plant ; and expressing the coding
region of
interest.
[0021 J The present invention also provides an isolated nucleotide sequence
comprising the nucleic acid sequence defined by SEQ ID N0:22, a nucleotide . '
sequence that hybridizes to the nucleic acid sequence of SEQ ID N0:22, or a
nucleotide sequence that hybridizes to a cor~.pliment of the nucleotide
sequence of
SEQ ID N0:22, wherein hybridi?ation condition is selected from the group
c~nsisting
of
hybridizing overnight in a solution comprising 7% SDS, O.SM NaP04 buffer
at pH 7.2, and 10 mM EDTA at 65°C and washing fbr one hour at
60°C in a solution
comprising 0.1 X SSC and 0.1% SDS;
hybridizing overnight in a solution comprising 7% SDS, O.SM NaP04 buffer
at pH 7.2, and 10 mM EDTA at 65°C and washing for one hour at
65°C in a solution
comprising 2 X SSC and 0.1% SDS; and
hybridizing overnight in a solution comprising 4 X SSC of 65°C and
washing
one hour in 0.1 X SSC at 65°C, and
wherein the nucleotide sequence exhibits regulatory element activity and is
capable of
mediating transcriptional efficiency of a transcript encoding a gene of
interest.
[0022] The present invention also pertains to a chimeric construct comprising
the
isolated nucleotide sequence as just described operatively linked with a
coding region
of interest. Furthermore, the present invention provides a method of
expressing a
coding region of interest within a plant comprising introducing the chimeric
construct
6
CA 02507563 2005-05-13
just defined, into a plant, and expressing the coding region of interest. The
invention
also includes a plant comprising the chimeric construct, a seed comprising the
chimeric construct, a plant cell comprising the chimeric construct. The plant,
seed or
plant cell may be selected from the group consisting of: a monocot plant, a
dicot plant, ,
a gymnosperm, an angiosperm, a hardwood tree, a softwood treep a cereal plant,
wheat, barley, oat, corn, tobacco, Brassica, soybean, pea, alfalfa, potato,
ginseng,
Arabidopsis, a peach, a plum and a spruce.
[0023] The present invention pertains to the isolated nucleotide sequence a
just
defined, wherein the nucleotide sequence is defined by SEQ ID NO:1, 5,~6, 7,8,
9, 21
Or 22, a nucleic acid sequence that hybridizes to the nucleotide sequence of
SEQ ID
NO:1, 5, 6, 7, 8, 9, 21 or 22, or a nucleic acid sequence that hybridizes to a
compliment of the nucleotide sequence of SEQ ID,NO:1, 5, 6, 7, 8, 9, 21_ or
22.
[0024] The present invention also pertains to a chimeric construct comprising
the
isolated nucleotide sequence as just described operatively linked with a
coding region
of interest. Furthermore, the present invention provides a method of
expressing a
coding region of interest within a plant comprising introducing the chirr~eric
construct
just defined, into a plant, and expressing the coding region of interest. The
invention
also includes a plant comprising the chimeric construct, a seed comprising the
chimeric construct, a plant cell comprising the chimeric construct. The plant,
seed or
plant cell may be selected from the group consisting of: a monocot plant, a
dicot plant,
a gymnosperm, an angiosperm, a hardwood tree, a softwood tree; a cereal plant,
wheat, barley, oat, corn, tobacco, Brassica, soybean, pea, alfalfa, potato,
ginseng,
Arabidopsis, a peach, a plum and a spruce.
[0025] The present invention also provides an isolated nucleotide sequence
comprising the nucleic acid sequence defined by nucleotides 1660-1875 of SEQ
ID
NO:1, a nucleotide sequence that hybridizes to nucleotides 1660-1875 of SEQ ID
NO:1, or a nucleotide sequence that hybridizes to a compliment of nucleotides
1660-
1875 of SEQ ID NO:l, wherein hybridization condition is 65°C over night
in 7%
7
CA 02507563 2005-05-13
SDS; O.SM NaP04; l OmM EDTA, followed by two washes at 50°C in 0.1
X SSC,
0.1 % SDS for 30 minutes each, wherein the nucleotide' sequence exhibits
regulatory
element activity and is capable of mediating transcriptional efficiency, of a
transcript
encoding a gene of interest.
[0026] The present invention also pertains to a chimeric construct comprising
the
isolated nucleotide sequence as just described operatively linked with.a
coding region '
of interest. Furthermore, the present invention provides a method of
expressing a ' '. , ,
coding region of interest within a plant comprising introducing the chimeric
construct
just defined, into a plant, and expressing the coding region of interest. The
invention
also includes a plant comprising the c_h_imeric construct, a seed comprising
the
chim2ric canstruct, a plant cell comprising the chimeric construct. The plant,
seed or
plant cell may be selected from tle group consisting of: a monacot plant, a
divot pla:~t,
a gymnosperm, an angiosperm, a hardwood tree, a softwood tree, a cereal plant,
wheat, barley, oat, corn, tobacco, Brassica, soybean, pea, alfalfa, potato,
ginseng,
Arabidopsis, a peach, a plum and a spruce. '
[0027] The present invention pertains to the isolated nucleotide s,:quer~c;,
just defined,
wherein the nucleotide sequence is defined by nucleotides 1660-1992 of SEQ ID
NO:1.
[0028] The present invention also pertains to a chimeric construct comprising
the
isolated nucleotide sequence as just described operatively linked with a
coding region
of interest. Furthermore, the present invention provides a method of
expressing a
coding region of interest within a plant comprising introducing the chimeric
construct
just defined, into a plant, and expressing the coding region of interest. The
invention
also includes a plant comprising the chimeric construct, a seed comprising the
chimeric construct, a plant cell comprising the chimeric construct. The plant,
seed or
plant cell may be selected from the group consisting o~ a monocot plant, a
divot plant,
a gymnosperm, an angiosperm, a hardwood tree, a softwood tree, a cereal plant,
8
CA 02507563 2005-05-13
wheat, barley, oat, corn, tobacco, Brassica, soybean; pea, alfalfa, potato,
ginseng,
Arabidopsis, a peach, a plum and a spruce. ' ~ ~ , ,
[0029] The lpresent invention relates to an isolated nucleotide sequence
comprising
the nucleic acid sequence defined by nucleotides 2091-2170 of SEQ ID NO:l, a
nucleotide sequence that hybridizes to nucleotides 2091-2170 of SEQ ID NO:1;
or a
nucleotide sequence that hybridizes to a compliment of nucleotides 209.1-2170
of
SEQ ID NO:1, wherein hybridization condition is 65°C over night in 7%
SDS;.O.SM
NaP04; lOrriM EDTA, followed by two washes at 5f°C in 0.1 X SSC, 0.1%
SDS for
30 minutes each, wherein the nucleotide sequence exhibits regulatory element
activity
and is capable of mediating transcriptional efficiency of a transcript
encoding a gene
of iraerest.
[0030] The present invention also pertains to a chimeric construct comprising
the
isolated nucleotide sequence as just described operatively linked with a
coding region
of interest. Furthermore, the present invention provides a method of
expressing a 1
coding region of interest within a plant comprising 'introducing the chimeric
construct
just defined, into a plant, and exprPssir;g the coding region of interest. The
invention
also includes a plant comprising the chimeric construct, a seed comprising the
chimeric construct, a plant cell comprising the chimeric construct. The plant,
seed or
plant cell may be selected from the group consisting of: a monocot plant, a
dicot plant;
a gymnosperm, an angiosperm, a hardwood tree, a softwood tree, a cereal plant,
wheat, barley, oat, corn, tobacco, Brassica, soybean, pea, alfalfa, potato,
ginseng,
Arabidopsis, a peach, a plum and a spruce.
[0031] The present invention also pertains to the isolated nucleotide sequence
as just
described, wherein the nucleotide sequence is defined by nucleotides 1660-2224
of
SEQ ID NO:I, 1723-2224 of SEQ ID NO:1, 415-2224 of SEQ ID NO:1, 1040-2224
of SEQ ID NO:1, 1370-2224 of SEQ ID NO:1, 2084-2224 of SEQ ID NO:1, or 2042-
2224 of SEQ ID NO:1.
9
CA 02507563 2005-05-13
[0032] The present invention also pertains to a chimeric construct comprising
the
isolated nucleotide sequence as just described operatively linked with a
coding region
of interest. furthermore, the present invention provides a method of
expressing a
' ,
coding region of interest within a plant comprising introducing the ehimeric
construct
just defined, into a plant, and expressing the ,coding region of interest.
'the invention , '
also includes a plant comprising the chimeric construct, a seed comprising the
chimeric construct, a plant cell comprising the chimeric construct. The plant,
seed or
' ~ ' ~ i I
plant cell may be selected from the group consisting of: ,a monocot plant, a
dicot plant;
a gyrranosperm, an angiosperm, a hardwood tree, a. Softwood tree, a cereal
plant,
wllPat_ barlPV, oat. corr~~ tnb~r.rn5 ~rcrc~trn etwhPan~ YP~~ alf~lfa~ ~n~a~v~
gin~nvrig~ ,'
Arabia'opsis, a peach, a plum and a spruce. , ',
~. ~ ,
[00331 The present invention provides an isolated nucleotide sequence
co~nprising the
nucleic acid sequence defined by nucleotides 1875-1992 of SEQ ID NO:1, a
nucleotide~sequence that hybridizes to nucleotides' 1875-1992 of SEQ ID NO:1,
or a
nucleotide sequence that hybridizes to a compliment of nucleotides 1875-1'992
of
$F.Q ID NQ:I w_h_Prei__n_ ltylJrjrli_Zatinn ~Cnditinn ~~~ HS°lv' C:'er
night 1T'. ~~~ SDS; ~.~l~i
NaPO~; IOmM EDTA, followed by two mashes at 50°O in 0.1 X SSC, 0.1%
SDS for
30 minutes each, wherein the nucleotide sequence exhibits regulatory element
activity
and is capable of mediating transcriptional efficiency of a transcript
encoding a gene
of interest.
[0034] The present invention also pertains to a chimeric construct comprising
the
isolated nucleotide sequence as just described operatively linked with a
coding region
of interest. Furthermore, the present invention provides a method of
expressing a
coding region of interest within a plant comprising introducing the chimeric
construct
just defined, into a plant, and expressing the coding region of interest. The
invention
also includes a plant comprising the chimeric construct, a seed comprising the
chimeric construct, a plant cell comprising the chimeric construct. The plant,
seed or
plant cell may be selected from the group consisting of: a monocot plant, a
dieot plant,
a gymnosperm, an angiosperm, a hardwood tree, a softwood tree, a cereal plant,
CA 02507563 2005-05-13
wheat, barley, oat, corn, tobacco, Brassica, soybean,~pea, alfalfa, potato,
ginseng,
Arabidopsis, a peach, a plum and a spruce. ' ~ . , ~ ,
[0035] The present invention pertains to an isolated nucleotide sequence as
just
described, wherein the nucleotide sequence is defined by nucleotides 1875-2084
of
SEQ ID NO:l . Furthermore, the nucleotide sequence defined by nucleotides 1875-
2084 of SEQ ID NO:1 may be present in tandem.
[0036] The present invention also pertains to a chirr~eric construct
comprising the
isolated nucleotide sequence as just described operatively linked with~a
coding region
~;,~+oroc.+ T.".-+1-,ovw",.-A +l.,o ov.+' +:,... '.a,.~. +1....,.7 1 ~..-~..
V1 111WLWV. 1 UW 11v1111V11.r, 111v pluJV111. 111Ve11L1V11 ~J1V VllilJd Q
111~L11VU 1J1 V11f.11ev1J111~,' Q
coding region of interest within a plant comprisir_g introducing the chimeric
construct
just defined, into a plant, and expressing the coding region of interest. The
invention
also includes a plant comprising the chimeric construct, a seed comprising the
chimeric construct, a plant cell comprising the chimeric construct. The plant,
seed or
plant cell may be selected from the group consisting of: a monocot plant; a
dicot plant,
a gymnosperm, an angiosperm, a hardwood tree, a softwood tree, a cereal plant,
wheat, barley, oat, corn, tobacco, Brassica, soybean, pea, alfalfa, potato,
ginseng,
Arabidopsis, a peach, a plum and a spruce.
(0037] The present invention also provides an isolated nucleotide sequence
comprising the nucleic acid sequence defined by nucleotides 1-1660 of SEQ ID
NO:l,
a nucleotide sequence that hybridizes to nucleotides 1875-1660 of SEQ ID~NO:1,
or a
nucleotide sequence that hybridizes to a compliment of nucleotides 1-1660 of
SEQ ID
NO:1, wherein hybridization condition is 65°C over night in 7% SDS;
O.SM NaP04;
lOmM EDTA, followed by two washes at 50°C in 0.1 X SSC, 0.1% SDS for 30
minutes each, wherein the nucleotide sequence exhibits regulatory element
activity
and is capable of mediating transcriptional efficiency of a transcript
encoding a gene
of interest.
[0038] The present invention also pertains to a chimeric construct comprising
the
isolated nucleotide sequence as just described operatively linked with a
coding region
11
CA 02507563 2005-05-13
' ,
n , i S
of interest. Furthermore, the present invention provides a method of
expressing. a
,
coding region of interest within a plant comprising introducing the chimeric
construct
just defined, into a plant, and expressing the coding region of interest. The
invention
a ,
also includes a plant comprising the chimeric construct, a seed comprising the
'
chimeric construct, a plant cell comprising tl~e chimeri~ construct. The
plant,' seed or , '
plant cell may be selected from the group consisting of: a monocot plant, a
dicot plant,
a gymnosperm, an angiosperm, a hardwood tree, ~a softwood tree, a cereal
plant,
~~ ~ , n ,
wheat, barley, oat, corn, tobacco, Brassica, soybean, pea, alfalfa, potato,
ginseng, ~ ''
Arabi,dopsis, a peach, a plum and a spruce.
, . ,.
[Ot~39] The present invention provides an isolated nucleotide sequence
comprising the
following nucleic acid sequence: '
TTATAATTAC AAAATTGATT MTAGTWYYTT TAATTTAATR YTTWTACATT ,'
ATTAATTAAY TTAGHWSTTT,YAATTYDTTT TCARAAAYYA TTTTACTATK:
KTT(T/-)RT AAAAWMAAAR GGR~AAARTG GYTATTTAAA TACYAAC(M/-)
. ,
CTATTTYATT TCAATTWTAR CCTAAAATCA,R(M/-)CCC(C/-) ARTTARCGCC ~
(W/-)(A/-) (T/-)(T/-) (Y/-)(C/-) (A/-)(A/-) (A/-)(T/-)
(T/-)(C/-) AAAYGGBMYA KCCCARTTCC TAAA(A/-)Y RACYCDCYCC
TAACCC (K/-) (C/-) (T/-) (T/-) (W/-) (T/-) (C/-) (C/-) (A/-)
(A/-)(C/-) (C/-)(C/-) RCCCKRTTYC CYCTTTTGAT CCAGGYYGTT
GATCATTTTG ATCAACGVCC ARAATTTCCC CYTTYC(Y/-) (K/-)TTTT
TMATTCCCAA ACACC(S/-) CCYAAMYYTA TCCCRTTTCT CACCAACCGC
CAGATMT(R/-)(W/-)(A/-)(T/-)CCTCT TATCTCTCAA ACTCTCTCGA
ACCTTCCCCT AACCCTAGCA GCCTCTCATC ATCCTCACCT CAAAACCCAC
CGGMMWMCAT GGCYTCTMRA G(S/-)(M/-)(K/-)(Y/-) (G/-)(R/-)
(W/-)(M/-) (M/-)(C/-) (C/-)(K/-) (K/-)(R/-1 (T/-)(R/-)
(S/-)(T/-) (C/-)(A/-)( S/-)(Y/-) YCCYYD(T/-)(G/-)(Y/-)
(N/-)(M/-) (T/-)(T/-) (A/-)~
a nucleotide sequence that hybridizes to the nucleic acid sequence, or a
nucleotide
12
CA 02507563 2005-05-13
sequence that hybridizes to a compliment of the nucleotide sequence, where R
is G or
.
A;YisTqrC;MisAorC;KisGorT;SisGorC;WisAorT;BisGorCorT;
DisAorGorT;HisAorCorT;andNisAorCorTorG,andwherein
hybridization is selected from the group consisting of : '
hybridizing overnight in a solution comprising 7% SDS, 0.5M NaP04 buffer
at pH 7.2, and 10 mM EDTA at 65°C and washing for one hour at
60~°C~in a solution
i , .
comprising 0.1 X SSC and 0.1% SDS; ~ '
. ,
~~ '
.
hybridizing overnight in a solution cpmprising 7Q/o SDS, O.SM NaP04 .buffer ,
at pH 7.2, and 10 mM EDTA at.65°C and washing for one hour at
65°C in a solution
comprising 2 X SSC and 0.1% SDS; and
hybridizing overnight in a solution comprising 4 X SSC at 65°C and
washing ,
one hour in 0.1 X SSC at 65°C, and
wherein the nucleotide sequence exhibits regulatory element activity and is
capable of
a:..+:~~+_..~,.,._:_+:.._...~ rr:..:~_.._. r..4_........_..:._4 .~:_._
r:._+._...._.+
Wcmamy uaiiaciiriiviiai cmncmy W a uamcty 2iiCOum~ a gene of mucicm.
[0040] The present invention also pertains to a chimeric construct comprising
the
isolated nucleotide sequence as just described operatively linked with a
coding region
of interest. Furthermore, the present invention provides a method of
expressing a
coding region of interest within a plant comprising introducing the chimeric
construct
just defined, into a plant, and expressing the coding region of interest. The
invention
also includes a plant comprising the chimeric construct, a seed comprising the
chimeric construct, a plant cell comprising the chimeric construct. The plant,
seed or
plant cell may be selected from the group consisting of: a monocot plant, a
dicot plant,
a gymnosperm, an angiosperm, a hardwood tree, a softwood tree, a cereal plant,
wheat, barley, oat, corn, tobacco, Brassica, soybean, pea, alfalfa, potato,
ginseng,
Arabidopsis, a peach, a plum and a spruce.
13
CA 02507563 2005-05-13
[0041] The present invention discloses transgenic plants generated by tagging
with a
promoterless GUS (13-glucuronidase) T-DNA vector and the isolation and . ,
characterization of a regulatory element identified usirig~this protocol.
Cloning and
characterization of this insertion site uncovered a unique regulatory element
not
conserved among related species. The novel constitutive regulatory element is
.
expressed in tissues throughout a plant and across a broad range of plant
species. The
novel constitutive regulatory element as described herein comprises additional
regulatory elements, and is a member of a large family of repetitive elements
that also
exhibit regulatory element activity. Therefore, the present invention also
describes
nnP nr mnrP than yP _n_pvP.l rPaapla_tnr~r PlemPnt ayd itc hnmning~,
FprthPrm~rP~ n~~rPl
non-translated 5' sequences have been identified within the regulatory element
that
function as post transcriptional regulatory elements.
[0042] This summary of the invention does not necessarily describe all
features of the
invention. ,
BRIEF DESCRIPTION OF THE DRAWINGS
[0043] These and other featares of the invention will become more apparent
from the
following description in which reference is made to the appended drawings
wherein:
[0044] FIGURE 1 shows the constitutive expression of GUS in all tissues of
plant
T1275, including leaf segments (a), stem cross-sections (b), roots (c), flower
cross-
sections (d), ovary cross-sections (e), immature embryos (fJ, mature embryos
(g), and
seed cross-sections (h).
[0045] FIGURE 2 shows GUS specific activity within a variety of tissues
throughout
the plant T 1275, including leaf (L), stem (S), root (R), anther (A), petal
(P), ovary (O),
sepal (Se), seeds 10 days post anthesis (Sl), and seeds, 20 days post anthesis
(S2).
[0046] FIGURE 3 is the Southern blot analysis of Eco RI digested T1275 DNA
with
a GUS gene coding region probe (lane 1) and a nptll gene coding region probe
(lane 2)
revealing a single T-DNA insertion site in plant T1275.
14
CA 02507563 2005-05-13
. n' , S
[0047] FIGURE 4 shows the cloned GUS gene fusion from pT1275. Figure 4 (A)
shows a restriction map of the plant DNA sequence fused with GUS. Figure 4(B)
.
' ,
shows the restriction map of the plant DNA. The arrow indicates the GUS mRNA
~'
i
start site within the plant DNA sequence. '
' ,
[0048] FIGURE 5 shows deletion constructs of the T1275 (tCUP) regulatory , '
element, and several results obtained with these constructs. Figure ~ (A)
shows the ' '
restriction map of the plant DNA of pT1275 upstteam firom the GUS insertion
site. ' ': , ,,
Figure 5 (B) shows further deletion constructs of -62-GUS=nos, -12-GUS-nos, -
62(-
tsr)-GUS-nos and +30-GUS-nos, relative to ,-197-GUS-nos (see Figure 5 (A)).
'Figure
(C) shows the 5' endpoints of each construct as indicated by the restriction
endonuclease site, relative to the full l~ngtr.,T1275 (tCUI~) regulatory
element, tr~e
arrow indicates the transcriational start sites Plant DNA is indicated by the
solid ~i~e,
the promoterless GUS-nos gene is indicated by the open box and the shaded box
,
indicates the region coding for the amino terminal' peptide fused to, GUS. The
XbaI ,
fragment in pT1275 was subcloned to create pT1275-GUS-nos. Deletion constructs
'
:~ ,
trmc_a_ted at t_he ,SphI, _ _ _Pstl .c YT~ RctYT~ and rJrwl sites were also
subcloned to create =
1639-GUS-nos, -1304-GUS-nos, -684-GUS-nos, -394-GUS-nos, and -197-GUS-nos,
respectively. Figure 5 (D) shows modified constructs of the T1275 regulatory
elements. T1275 is indicated by the open box, the CaMV35S promoter element is
indicated by the black box. The activity of these constructs is also
indicated. GUS
activity was determined in tobacco leaves following transient expression using
microparticle bombardment. TA30-GUS: a TATATAA element was inserted into the
-30 position of -62-GUS; TA35S-GUS: the -62 to -20 fragment of -62-GUS was
substituted with the -46 to -20 fragment of the 35S promoter; GCC-62-GUS: a
GCC
box was fused with -62-GUS; DRA2-GUS: the -197 to -62 fragment was repeated;
BST2-GUS: the -394 to -62 fragment was repeated; -46-35S: 35S minimal
promoter;
DRAI-355: the -197 to -62 fragment of T1275 was fused with -46-355; BSTI-355:
the
-394 to -62 fragment of T1275 was fused with -46-355; BST2-355: two copies of
the
-394 to -62 fragment of T1275 were fused with -46-355. Figure 5 (E) shows
CA 02507563 2005-05-13
constructs of the -197 to -62 fragment fused with the 35S minimal promoter. -
46-355:
35S minimal promoter; DRAI-355: the -197 to -62~frxgment of T1275 was fused
with
-46-35S; DRA1R-355: the -197 to -62 fragment ofT1275 was fused with -46-35S in
a'
reversed orientation; DRA2-355: two copies of the -197 to -62 fragment of
T1275
were fused with -46-355. Figure 5 (F) shows GUS specific activity of
transgenic
Arabidopsis plants. Leaf tissues from Arabidopsis plants transformed with -47-
355,
DRAM -3 5 S, DRA 1 R-3 5 S and DRA2-3 5 S constructs were used for GUS assay.
Figure 5 (G) shows the constitutive expression of GUS in Arabidopsis plants
transformed with DRA1-355. From top to bottom (i.e. Figures SG(i), SG(ii) and
SG(iii), respectively): flower, silque and seedling. 'Figure 5 (H) shows the
schematic
diagram of the chimerical constructs. The numbers on the top indicate deletion
end
points relative tc tr~e trarscripticr~ ir~itiatior~ site (+1) cf the tCUP. The
position of
transcription start site is i~?dicated by an arrow. The dot line indicates the
sequence
been deleted. These constructs include (see Figure 5 (B) for more
information): "-62"
(-62T1275-GUS-nosy; "-12" (-12T1275-GUS-nosy; "-62-tsr" (-61(-tsr)-GUS-nosy;
TA30 (sequence -30 to -24 of T1275 is replaced with TATATAA); GC'C-62
(addition
of (~'.C~'(''-hnx cPn»anrPCl T11a111'P ~ !Tl chn~arc tha rPlativa artivitv of
tha r~nnctr"~tc
..._ ... ~ ~ _ .._.1..._........~. s .b...~ ., ~. ~~~ "..., .... ...,.
....,...,.... .....~.....) ...~ ...... ...~..,m......w
outlined in Figure 5 (Hl within tomato protoplasts. Each value _represents the
average
of four independent experiments. Error bars indicate SE values. Figure 5 (~
shows
schematic diagrams of the 5' deletions chimerical constructs. -394(2X)-GUS and
-
197(2X)-GUS are the two constructs to test the effect of reiteration of the
tCUP
upstream regions (-394 to -62 and -197 to -62) on promoter activity. The
numbers on
the top indicate deletion end points relative to the transcription initiation
site (+1) of
the tCUP promoter. Figure 5 (K), shows the average GUS specific activity (pmol
MU /min/ mg protein) in transgenic Arabidopsis plants containing constructs
shown
in Figure 5 (J). 15-20 independent transgenic plants were tested for each
construct.
Figure 5 (L) shows the schematic diagram of chimerical constructs to study the
effect
of the tCUP upstream region -197 to -62 on -46 minimal CaMV 35S promoter
activity. The numbers on the top indicate deletion end points relative to the
transcription initiation site. Open-boxes represent the tCUP sequence and
filled-boxes
16
CA 02507563 2005-05-13
represent the CaMV 35S promoter sequence. Figure 5 (M) shows the verage GUS
specific activity (pmol MU /min/ mg protein) in trans~enic Arabidopsis plants
,
containing constructs shown in A. 15-20 independent transgenic plants were
tested for
each construct. ~ '
[0049] FIGURE 6 shows the GUS specific activity, mRNA, and protein levels in ,
~ '
leaves of individual, regenerated, greenhouse-grown transgenic tobaGCO plants
' '
containing T1275-GUS-nos (T plants), or 35S-BUS-nos (S plants). Figure 6 (A) '
'; , ,,
shows the levels of GUS expression in leaves from~r~ndomly selected plants
,
containing either T1275-GUS-nos (left-hand side)'or 35S-GUS-nos
(right=hand'side).
Figure 6 (B) shows the level of accumulated G[IS mRNA measured by RNase
protection assay and den sito:retry of autoradiogra.-ns in l;,aves frorr~ the
same
randomly selected plants containing either T1275-C=T,JS-nos (left-hand side)
or 35,5-
GUS-nos (right-hand side). Figure 6 (C) shows a Western blot of GUS fusion
protein
obtained from T1275-GUS-nos and 35S-GUS-rios plants. Leaf extracts were
equally
loaded onto gels and GUS was detected using anti=GUS antibodies. The molecular-
'
wei_aht marlcerc are indicated nn the riaht_hanwl Bide nfthP ~rPl~
,~ntrangfnrme~l ~rnntrnl
,
(SR1) and GUS produced in E. coli (Ec).
[0050] FIGURE 7 shows deletion and insertion constructs of the 5' untranslated
leader region of T1275 regulatory element and construction of transformation
vectors.
The constructs are presented relative to T1275-GUS-nos or 35S-GUS-nos. The
arrow
indicates the transcriptional start site. Plant DNA is indicated by the solid
line labeled
T1275, the 35S regulatory region by the solid line labelled CaMV35S, the NdeI -
SmaI
region by a filled in box, the shaded box coding for the amino terminal
peptide, and
the promoterless GUS-nos gene is indicated by an open box. The deletion
construct
removing the NdeI - SmaI fragment of T1275-GUS-nos is identified as T1275-N-
GUS-nos. The NdeI - SmaI fragment from T1275-GUS-nos was also introduced into
35S-GUS-nos to produce 35S+N-Gus-nos.
17
CA 02507563 2005-05-13
[0051 ] FIGURE 8 shows the region surrounding the insertion site in
untransformed
plants, positions of various probes used for RNase protection assays,
and~results o~the
RNase protection assay. Figure 8 (A) shows a restriction map of the insertion
site
' and various probes used for the assay (IP: insertion point of GUS in
transformed
plants; * : that T 1275 probe ended at the BstYl site, not the IP; * * :
,probe 7 included
600bp of the T1275 plant sequence and 400 by of the GUS gene). Figure 8 (B) '
shows results of an RNase protection assay of RNA isolated from leaf (L), stem
(St), ,
root (R), flower bud (F) and developing seed (Se) tissues of tobacco
transformed with
T1275-GUS-nos (10 pg RNA) and untransformed tobacco (30 gg RNA). Undigested
probe (p)~ r_t~a negative control (-) ~a'nes and markers are indicated.
F~l~Tase
proteCtiO?'I - -- rr -asSays S11oW11 used a probe t0 det~~t S°nC~
tr~_n_cCriptg hAfy.~ACn a~,nyt -446
and +596 of T1275-GUS-nos or between about -446 to +169 of untransformed
tobacco. The protected fragment in transformed plants is about 596 by (upper
arrowhead) and, if present, accumulated transcripts initiated at this site in
untransformed plants are predicted to protect a fragment of about 169 by
(lower
arrowhead). Upper band in RNA-containing lanes was added to samples to
indicate
loss of sample during assay.
[0052] FIGURE 9 shows the levels of mRNA, as well as the ratio between GUS
specific activity and mRNA levels in leaves of individual, regenerated,
greenhouse-
grown transgenic plants containing T1275-GUS-nos (i.e. tCUP-GUS-nosy, or 35S-
GUS-nos constructs, with or without the Ndel Smal fragment (see Figure 7).
Figure
9 (A) shows the level of accumulated GUS mRNA measured by RNase protection
assay and densitometry of autoradiograms in leaves from the same randomly
selected
plants containing either T1275-GUS-nos, T1275-N-GUS-nos. Figure 9 (B) shows
the
level of accumulated GUS mRNA measured by RNase protection for 35S-GUS-nos or
35S+N-GUS-nos. Figure 9 (C) shows the ratio between GUS specific activity and
mRNA levels in leaves of individual, regenerated, greenhouse-grown transgenic
plants containing tCUP-GUS-nos, tCUP-N-GUS-nos, 35S-GUS-nos, or 35S+N-GUS-
nos constructs.
18
CA 02507563 2005-05-13
[0053] FIGURE 10 shows the maps of T1275-GUS-nos and T1275(ON)-GUS-nos.
Figure 10 (A) shows T1275-GUS-nos (also referred to as,tCUP-GUS-nas). Figuxe
(B) shows T1275(~N)-GUS-nos (also referred to as tCUPdelta-GUS-nosy. "0N",
(also referred to as "dN" or "deltaN") was created by changing the NdeI site
"a" in the
leader sequence of T1275-GUS-nos (Figure 10(A)) to a BgIII site ;'b" (see
Figure
10(B)) to eliminate the upstream ATG at nucleotides 2087-2089 of SEQ ID N0:2.
'A
Kozak consensus sequence "c" was constructed at the initiator MET codon and a
NcoI ,
site was added. The transcriptional start site, determined for T1275, is
indicated by
the arrow. '
X0054] FIGURE 11 shows constructs used for the transient expression via
particle
bombard~-ner~t of cam callus. Maps for 35S-GUS-nos, 35S (+NT)-GUS-nos, 35S
(~N)-
GUS-nos and 35S(+i)-GT".JS-nos are presented indicating the "N" region, ADT.-
il
intron, and the arrow indicates the transcriptional start site. Note that
35S(ON)-GUS-
nos is referred to as 35S+deltaN-dK-GUS-nos. Also shown are the associated, .
activities of the constructs in the callus expressed as a ratio of GUS to
luciferase
(control) activity.
[0055] FIGURE 12 shows maps of the constructs used for transient expression in
yeast. Shown are pYES-GUS-nos (also referred to as pYEGUS); pYES(+N)-GUS-nos
(also referred to as pYENGUS); pYES(ON)-GUS-nos (also referred to as
pYEdNGUS) and pYES(ONM)-GUS-nos (also referred to as pYEdNMGUS), which
lacks the Kozak consensus sequence.
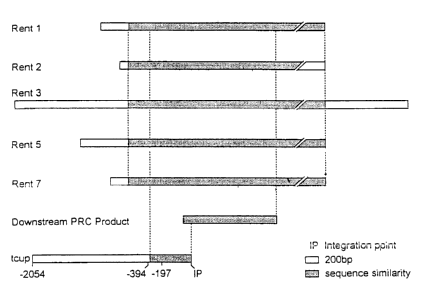
[0056] FIGURE 13 shows the sequence similarity between several members of the
RENT family of highly repetitive sequences. Figure 13 (A) shows a homology
tree of
an approximately 600 by fragment of RENT 1 (SEQ ID N0:5), RENT 2 (SEQ ID
N0:6), RENT 3 (SEQ ID N0:7), RENT 5 (SEQ ID N0:8), RENT 7 (SEQ ID N0:9)
and T1275 (tCUP; SEQ ID NO:1). Figure 13 (B) shows a graphic representation of
the sequence alignments between the different RENT clones and T1275 (tCUP).
Figure 13 (C) shows the actual sequence alignments of Figure 13 (B), where the
19
CA 02507563 2005-05-13
numbering above the sequences indicates the numbering relative to RENT 7 (SEQ
ID
N0:9), and the numbering below the sequences indicate the alignment of the
RENT
consensus sequence (SEQ ID N0:21 ) relative to the tCUP ~ sequence (SEQ ID
NO:1 ).
The consensus sequence relative to tCUP islpresented. Inserts within the RENT
consensus nucleotide sequence that are not present in tCUP, are indicated
above the
consensus sequence. Deletions in the nucleotide sequence in at least one
member of
the RENT family of nucleotide sequences that are not present in tCUP, are
indicated
as "-" above the consensus sequence. R is G or A; Y is fi or C; M is A~or C; K
is G or
T;S~sGorC;WisAorT;BisGorCorT;Dis.AlorGorT;HisAorCorT;and
N is A or C or T or G. Figur a 13(I3) shoves the RENT consensus sequence (SEQ
ID
-~ ~ P,.A d f y g o , ( f p
'._~O__! ), seP i.,a,,n r~~ , o m_r.~ . ~ (~'1 .nr GIPt~.g p'f ceqapnCC
rr,,~~gRn.~:l~n. ~:g'u: °
13(E) shows the nucleotide sequence for tCUP-RENT (SEQ ID N0:22) where
nucleotides 1-1723 comprise the nucleotide sequence of tCUP (SEQ ID NO:1), and
nucleotides from 1724 to 2224 comprise the RENT consensus sequence (SEQ ID
N0:21). '
[0057] FIGURE 14 shows the expression of a coding region of interest driven by
regulatory elements obtained from several members of the RENT family of highly
repetitive sequences. Figure 14 (A) shows the transient expression of
constructs
comprising a RENT regulatory element in operative association with GUS-nos,
and
the expression of these constructs in pea protoplasts. The constructs were
introduced
into pea protoplasts via electroporation (see methods for details). tCUP RENT
(PCR
fragment from 1772 of SEQ ID NO:1 fused to delta N); RENT 1 (SEQ ID NO:S),
RENT 2 (SEQ ID N0:6), RENT 3 (SEQ ID N0:7), RENT 5 (SEQ ID N0:8), RENT
7 (SEQ ID N0:9), 355-46 (35S minimal promoter. Figure 14 (B) shows
histochemical analysis of GUS expression in transgenic Arabidopsis plants
containing
-394tCUP-GUS construct. GUS gene was expressed in leaves, stems, flowers,
siliques
and roots of transgenic Arabidopsis plants.
CA 02507563 2005-05-13
DETAILED DESCRIPTION
~~ .
[0058] The present invention relates to regulatory elements obtained from a
plant.
This invention further relates to the use of one or more than one regulatory
element to
control the expression of exogenous DNAs of interest in a desired host.
[0059] The following description'is of a preferred embodiment. '
' ,
[0060] T-DNA tagging with a promoterless (3-glucuronidase (GUS) gene generated
several transgenic Nicotiana tabacum plants that expressed GUS activity. An
'example, which is not to be considered limiting in ahy manner, of transgeriic
plants
displaying expression of the promoterless reporter gene, includes a plant trat
expressed GUS in aii organs, T i 275 (see co-pending' patent applications U S
serial No.
08/593121, PCT/CA97/00064; and PCT/CA99/0057 which are incorporated by ~ ,
reference).
~ I T f"lnnina anrl rleletinn anal~rcig of the (":T TC f,,,5i~ng ;n tl~PSO.
nluntc rc»realerl that
... ._ Jv ~ v ivr a a m
one or more than one regulatory region was located in the plant DNA proximal
to the
GUS gene. In T1275, a regulatory region was identified within an XbaI - SmaI
fragment that exhibits constitutive activity in all organs, tissues and plants
tested. This
constitutive regulatory element, is referred to as T1275, or tCUP herein (SEQ
ID
NO's:l or 22), and comprises several other regulatory elements throughout the
sequence, and that exhibit regulatory region activity as defined herein, for
example:
- a minimal promoter region between DraI and NdeI sites (1875-2084 of SEQ
ID NO's:l and 22), also referred to as a,cqre promoter element; see Figure
SC "-197-GUS-nos", and Table 6;
- negative regulatory elements between 1040-1370 of SEQ ID NO's:l and 22
("-1304 to -684"; see Figures SJ and K, where activity obtained for
"tCUP" and "-684" are each above that of the activity obtained for "-
13 04");
21
CA 02507563 2005-05-13
,, I ,
s
- a transcriptional enhancer between BstYI and DraI sites (1660-1875 of SEQ
,ID NO's:l and 22), also referred to as a BstYI-DraI fragment; see Figure
I ,
SC e.g. "-394 GUS-nos", and Table, 6);~
a translational enhancer regulatory element between NdeI and SmaI site's , '
1 I~
(2084-2224 of SEQ ID NO's:l and 22) see Figures SB (+30-GUS-nosy, , ~ , '
Figure 7 (T1275-GUS-nos; 35S-GUS'-nos) and Tables 7-13.' This '
fragment is also referred to as "N" heiein. Also see Figure.l 1 (comp'are ' ';
. ,
'the activity of 35S+N-GUS=nos, comprising the INdeI-SmaI fragment, with I
t . ,
that of 35S-GUS-nos, lacking the~NdeI-SmaI~fragment). A shorten~'d I
fragment of N comprising nucleotides 2091-2170 of SEQ ID NO's:l and
' 22 (presented in SEQ ID X0:2; also referred to as dN, deltaN, tCUP '
delta), ~NM (a fragment that lacks a Kozak sequence; SEQ ID N0:4),, or a
fragment that comprises a Kozak sequence (Figure 10, SEQ ID N0:3) also
exhibit enhancer regulatory. element activity.
- an enhancer element between 1660-1992 of SEQ ID NO's:l and'22
(fragment between BstYI ("-394") and "-62"), see Figure SD (see Bstl-
GUS; Bstl-35S, and tandem fragments: Bst2-GUS, Bst2-35S);
- a transcriptional enhancer between 1875-1992 of SEQ ID NO's: l and 22
(fragment between Dral ("-197") and "-62"), see Figure SD (Dral-GUS;
Drat-GUS; Dral-355; Dra2-35S), and Figures SE-G (Dral-355; Dra2-
35S); and
- members of the RENT family exhibit greater than 75% sequence identity
with nucleotides 1724-2224 of SEQ ID NO:1, or more preferably, from
about 77% to 92 % sequence identity with nucleotides 1724-2224 of SEQ
ID NO:I (see Figures 13C, 13D and 14A. This region includes several of
the regulatory elements identified above including the minimal promoter
between DraI and NdeI sites (1875-2086 of SEQ ID NO:1) and the
22
CA 02507563 2005-05-13
translational enhancer between NdeI and, SmaI sites (2084-2224 of SEQ ID
NO:l). The consensus sequence for meinbers,of the RENT family (SF;Q
,, .
1D NO's: 21 and 22) is presented in Figures ~13(C) = 13(E). ,
[0062] Therefore, the present invention provides one or more thap one
regulatory
region obtained from T1275 (tCUP; SEQ ID NO's:l or 22), wherein the regulatory
region may comprise: .
- the'full length sequence of SEQ ID NO:1,'SEQ ID N0:21, or SEQ ID
NO:22; , ,
- a ptclP:otodA gPrniA~r;C tat ~ ~~~_rir?i~Cg t~ ~E(~1 ID i~_T!~~ 1 ~Fll 1TZ
1~T(1-21
Z --J-- .- , .a, vm~ aiv i v. a, yr
SEQ ID NO:22;
- a nucleotide sequence that hybridizes to the compliment of SEQ ID NO:1,
SEQ ID N0:21, or SEQ ID N0:22; , .
- a fragment of SEQ ID NO:1, SEQ ID N0:21 or SEQ ID N0:22, or
- a nucleotide sequence that hybridizes to a fragment of SEQ ID NO:1 SEQ
ID N0:21, or SEQ ID N0:22,
wherein the nucleotide sequence exhibits regulatory element activity, or is
capable of
mediating transcriptional efficiency of a transcript encoding a gene of
interest that is
operatively linked thereto.
[0063] By a nucleotide sequence exhibiting regulatory element activity it is
meant that
the nucleotide sequence, when operatively linked with a coding sequence of
interest,
regulates, modifies or mediates the expression of the coding sequence. For
example, a
nucleotide sequence exhibiting regulatory element activity may function as a
promoter, a core promoter, a constitutive regulatory element, a negative
element or
silencer (i.e. elements that decrease promoter activity), or a transcriptional
or
translational enhancer, thereby regulating, modifying or mediating expression
of a
23
CA 02507563 2005-05-13
I , s i
coding region of interest that may be operatively linked thereto.
Hybridization
condition ray be selected from the group consisting o~f: ' ,
- hybridizing overnight in a. solution' coinprisirig 7% SDS, O.SM NaP04 buffer
at pH 7.2, and 10 mM EDTA ax 65°C and washing for one hour at
60°C in a solution
comprising 0.1 X SSC and 0.1% SDS; ~ , '
,,
- hybridizing overnight in a solution comprising 7% SDS, O.S~M NaP04 buffer '
at pH 7.2, and 10 mM EDTA at 65°C and washing for one hour at
65°C.in a solution ' ' '~
comprising 2 X SSC and 0.1 % SDS; and , ' ~ '
. ,
- hybridizing overnight in a solution comprising, 4 X SSC'at 65°C and
washing one
hour i_r10.1_ X SSC at 65°C. ''
[0064] Furthermore, the present invention exemplifies the use of one or~more
probes,
for example but not limited to nucleotides 1660-2224 of SEQ ID NO:1 (BstYI-
SmaI
fragment), that may be used identify members of the RENT family of sequences
(see ,
Examples "RENT Repetitive Element from N. tabacum family of repetitive
elements''
in the Examples).
[0065] However, it is to be understood that other portions of the isolated
disclosed
regulatory elements within T1275 (tCUP) may also exhibit activities in
directing
organ specificity, tissue specificity, or a combination thereof, or temporal
activity, or
developmental activity, or a combination thereof, or other regulatory
attributes
including, negative regulatory elements, enhancer sequences, or post
transcriptional
regulatory elements, including sequences that affect stability of the
transcription or
initiation complexes or stability of the transcript. The full-length
nucleotide sequence
of the T1275 (tCUP) regulatory region is provided in SEQ ID NO:1. Nucleotide
sequences that exhibit from about 75% sequence identity with nucleotides from
about
1724 to 2224 of the T1274 regulatory region (SEQ ID NO:1), and that exhibit
regulatory element activity, are also disclosed. These nucleotide sequences
include
members of the RENT family of nucleotide sequences (see Figure 13C), and when
24
CA 02507563 2005-05-13
,
~, ,
operatively linked with a coding region of interest; drive the expression of
the coding
region of interest (see Figure 14(A)).
[0066] Thus, the present invention includes, but is not limited to one'or more
than one ; ,
regulatory element obtained fr4m plants that is capable, of conferring,
rr~ediating,
modifying, reducing, or enhancing expression upon a coding region of interest
~ , '
operatively linked therewith. Furthermore; the,present invention includes one
or more ' '.
than one regulatory element obtained from a plant that i,s capable of
mediating the ~ ~; , ,,
translational efficiency of a transcript produced frorri a coding region of
interest linked'
in operative association therewith.' It is to b~ understood that the
regulatory elements
of the present invention may also be used iri combination with other
regulatory '
elements, either cryptic or otherwise, 'such as promoters,'enhancers, or
fragments'
thereof, and the Like. . ,
[0067] Furthermore, the present invention provides an isolated plant
constitutive
regulatory element. This regulatory element may be characterized in that: ,
- it directs expression in a variety of plant tissues and organs, for example,
the
ovary, tl_ower, a_m__mature embryo, matprP emb_ryp~ cePd~ cterri~ 1_eaf _r~nt
and CpltprPd
tissues;
- it lacks a TATA box;
- it is not detected in untransformed soybean, potato, sunflower, Arabidopsis,
B. napus, or B. oleracea, corn, wheat, black spruce, by Southern analysis
under the
following conditions: 4XSSC at 65°C overnight (from 12-18 hours),
followed by
washing in 0.1 XS S C at 65 °C for an hour; and
- it is a member of a large family of repetitive elements (RENT).
[0068] The regulatory element described herein is a member of a large family
of
repetitive elements identified within the Nicotiana tabacum SR1 genome that
exhibits
greater than about 75%, and preferably from about 77% to about 90% sequence
2j
CA 02507563 2005-05-13
similarity to fragment of approximately 532 by of SEQ ID NO:1 (including
nucleotides 1724 to 2224; see Figures 13 (A) and (C); the,sequence.of tCUP in
Figure
13 (C) includes the tDNA portion of the T1275 sequence which comprise
nucleotides
635-667 of Figure 13(C)). This family of repetitive elements has been termed
RENT
(Repetitive Element Nicotiana~ tabacum). The approximately 532, by fragment of
SEQ
ID NO:1, and related nucleotide sequences as determined within the RENT family
(SEQ~ID NO's: 5 to 9), exhibit regulatory element activity and are capable of
directing
GUS expression in a range of plants. The RENT consensus sequence is provided
in
Figures 13(C)-(E) and in SEQ ID NO's:21 and 22.
[0069] This invention is also directed to a regulatory element that comprises
a
nucleotide sequence of at least 18 contiguous base pairs of SEQ ID NO'~s:l, 5,
6, 7, 8,
9, 21 or 22. Oligonucleotides of 18 by or more are useful in constructing
heterologous~
regulatory elements that comprise fragments of the regulatory element as
defined in
SEQ ID NO's:l, 5, 6, 7, 8, 9, 21, or 22. The use of such heterologous
regulatory
Wu..WeS iS 'v'v11 cStabiiS h':d ii1 ti3c iiiCtaIaALC. For example, fragments
of specific
elements within the 35S CaMV promoter have been duplicated or combined with
other promoter fragments to produce chimeric promoters with desired properties
(e.g.
U.S. 5,491,288; US 5,424,200; US 5,322,938; US 5,196,525; US 5,164,316).
Oligonucleotides of 18 bps or longer are useful as probes or PCR primers in
identifying or amplifying related DNA or RNA sequences in other tissues or
organisms. Furthermore, oligonucleotides of 18 bps or more are useful in
identifying
sequences homologous to those identified within SEQ ID NO's:l, 5 to 9, 21 or
22 for
example, but not limited to, the RENT family of elements, as described herein.
[0070] By "regulatory element" or "regulatory region", it is meant a portion
of nucleic
acid typically, but not always, upstream of a gene, and may be comprised of
either
DNA or RNA, or both DNA and RNA. The regulatory elements of the present
invention include those which are capable of mediating organ specificity, or
controlling developmental or temporal gene activation. Furthermore,
"regulatory
element" includes promoter elements, core promoter elements, elements that are
26
CA 02507563 2005-05-13 '
,,
s
inducible in response to an external stimulus, elements that are activated ~ '
,
constitutively, or elements that decrease or increase promoter activity such
as negative
regulatory elements or transcriptional enhancers, respectively. By a
nucleotide
,~ ,
sequence exhibiting regulatory element activity it is meant that the
nucleotide '
sequence when operatively linked with a coding sequence of interest functions
as a '
promoter, a core promoter, a constitutive regulatory element, a negative
element or
,, , ,
silencer (i.e. elements that decrease promoter activity), or a transcriptional
or
,,.
translational enhances. ' ' .
, ~ '
,
[0071 J ~By "operatively linked" it is meant that the particular sequences;
for .~Xample
a regulatory element and a coding region of interest, interact either directly
or
indi_reCtly to Carry apt an i__r~tP_n_rlPra yrtinn gyCh aS mediation or
I:~odulatior~ 0'f
gene expression. The interaction of operatively linked sequences may, for
exar.~ple,
be mediated by proteins that interact with the operatively linked sequences. I
~ ,
[0072) Regulatory elements as used herein,~also includes elements that are
active
_ , , ,
following transcription initiation or transcription, for example, regulatory
dements ' '
that modulate gene expression such as translational and transcriptional
enhancers,
translational and transcriptional repressors, and mRNA stability or
instability
determinants. In the context of this disclosure, the term "regulatory element"
also
refers to a sequence of DNA, usually, but not always, upstream (5') to the
coding
sequence of a structural gene, which includes sequences which control the
expression
of the coding region by providing the recognition for RNA polymerise and/or
other
factors required for transcription to start at a particular site. An example
of a
regulatory element that provides for the recognition for RNA polymerise or
other
transcriptional factors to ensure initiation at a particular site is a
promoter element. A
promoter element comprises a core promoter element, responsible for the
initiation of
transcription, as well as other regulatory elements (as listed above) that
modify gene
expression. It is to be understood that nucleotide sequences, located within
introns, or
3' of the coding region sequence may also contribute to the regulation of
expression of
a coding region of interest. A regulatory element may also include those
elements
27
CA 02507563 2005-05-13
located downstream (3') to the site of transcription initiation, or within
transcribed
regions, or both. In the context of the present invention a,post-
transcriptional ,
regulatory element may include elements that are active~follovving
transcription
initiation, for example translational and transcriptional enhancers,
translational and
transcriptional repressors, and mRNA stability determinants. ,
[0073,] The regulatory elements, or fragments thereof, of the present
invention may be
operatively associated (operatively linked) with heterologous regulatory
elements or
promoters in order to modulate the activity of the, heterologous
regulatory~element.
such modulation includes enhancing or repressing transcriptional activity of
the .
heterologous regulatory element, modulating post-transcriptional events, or
both
enhancing or repressing transcriptional activity of the heterologous
regulatory element
and modulating post-transcriptional events. For example, one or more
regulatory
elements, or fragments thereof, of the present invention may be operatively
associated
with constitutive, inducible, tissue specific promoters or fragment thereof,
or , ,
iagW2utS of rc~uiatGly GiG111Gilts, for exdir~pie, bui not limited to TATA or
il4
sequences may be operatively associated with the regulatory elements of the
present
invention, to modulate the activity of such promoters within plant, insect,
fungi,
bacterial, yeast, or animal cells.
[0074] T here are generally two types of promoters, inducible and constitutive
promoters. An inducible promoter is a promoter that is capable of directly or
indirectly activating transcription of one or more DNA sequences or genes in
response
to an inducer. In the absence of an inducer the DNA sequences or genes will
not be
transcribed. Typically the protein factor that binds specifically to an
inducible
promoter to activate transcription is present in an inactive form which is
then directly
or indirectly converted to the active form by the inducer. The inducer can be
a
chemical agent such as a protein, metabolite, growth regulator, herbicide or
phenolic
compound or a physiological stress imposed directly by heat, cold, salt, or
toxic
elements or indirectly through the action of a pathogen or disease agent such
as a
virus. A plant cell containing an inducible promoter may be exposed to an
inducer by
28
CA 02507563 2005-05-13
externally applying the inducer to the cell or plant such as by spraying,
watering,
heating or similar methods.
[4075] A constitutive promoter directs the expression of a gene throughout,the
various parts of a plant and continuously throughout plant development,
Examples of
known constitutive promoters include those associated with the CaMV 35S
transcript.
(Odell et al., 1985, Nature, 313: 810-812); the rice actin 1 (Zhang et ~1,
1991, Plant
Cell, 3: 1155-1165) and triosephosphate isomeras'e 1 (Xu et al, 1994, Plant
Physiol.
106: 459-467) genes, the maize ubiquitin 1 gene (Cornejo et al, 1993, Plant
Mol. Biol.
29: 637=646), the Arabidopsis ubiquitin 1 anc~ 6 genes (Holtorf et al, 1995,
Plarit Mol.
Biol. 29: 637-646), and the tobacco translationa~ iritiation factor 4A gene
(Mandel et
al, 1995 Pla:Zt I~ToI. Biol. 29: 995-1 CC-~+). The present invention is
directed to a DNA
sequence which contains a regulatory element capable, of directing the
expression, of a
gene. Preferably the regulatory elemerit is a constitutive regulatory element
isolated
from N. tabacum. '
[0076] The term "constitutive" as used herein does not necessarily indicate
that a gene
is expressed at the same level in all cell types, but that the gene is
expressed in a wide
range of cell types, although some variation in abundance is often observed.
[0077] An example, which is not to be considered limiting in any manner, of a
regulatory element of the present invention includes a constitutive regulatory
element
obtained from the plant T1275, as described herein and analogues or fragments
thereof, or a nucleic acid fragment localized between Xbal - Smal, as
identified by the
restriction map of Figure 4 (B) or a fragment thereof. Furthermore, the
regulatory
element may be defined as a nucleic acid fragment localized between XbaI -
SmaI as
identified by the restriction map of Figure 5 (C) or a fragment thereof. The
regulatory
element may also be defined by a nucleotide sequence comprising at least an 18
by
fragment of the regulatory region defined in SEQ ID NO's:l, 5, 6, 7, 8, 9, 21
or 22
The regulatory element may also be defined by a nucleic acid comprising from
about
70%, preferably greater than about 75%, nucleotide sequence similarity to the
29
CA 02507563 2005-05-13
nucleotide sequence of SEQ ID NO's: l , 5, 6, 7, 8, 9; 21 or 22 or a fragment
thereof, or
by a nucleic acid substantially homologous to the nucleotide sequence of SEQ
ID ,
NO's:l, 5, 6, 7, 8, 9, 21 or 22 or a fragment thereof, wherein the nucleic
acid exhibits
regulatory element activity.
[0078] Another regulatory element of the present invention includes, but is
not limited
to, a post-transcriptional or translational enhancer regulatory element
localized
between NdeI - SmaI (see Figures S (A), (B) or (C), Figure 7, and Figure 11),
or the
post-transcriptional or translational enhancer regulatory element may comprise
the
nucleotide sequence as defined by nucleotides 2084 -2224 of SEQ ID NO' 1 or an
.
analog thereof, or the element may comprise 70% similarity to the nucleotide
sequence of nucleotides 2084-2224 of SEQ ID i~TO:1 (i.e. a portion of the NdeI-
Smal
fragment from Nd~I to the integration point of T1275 at nucleotide 2224).
[0079] Furthermore, other regulatory elements of the present invention include
negative regulatory elements (for example located w',thi_n_ an XhnT-RctYT
fragment as
defined by Figure 5 (C), and described in more detail below), a
transcriptional
enhancer localized within the BstYI-DraI fragment of Figure 5 (C), a core
regulatory
element located within the DraI-NdeI fragment of Figure 5 (C), or a regulatory
element or post-transcriptional element downstream of the transcriptional
start site.
[0080] A further regulatory element of the present invention includes an
enhancer
element within the -394 to -62 fragment of T1275 (nucleotides 1660 to 1992 of
SEQ
ID NO:1). This fragment may also be duplicated and fused to a regulatory
region, for
example a core promoter, producing an increase in the activity of the
regulatory region
(see Figure 5 (D)). A portion of the -394 to -62 fragment of T1275 (tCUP),
from
nucleotides 1724-1992 of SEQ ID NO:1 or 22 exhibits substantial homology with
other members of the RENT family of repetitive sequences (Figures 13 (A) -
(C)).
The homologous fragment present within the RENT family of sequences also
exhibit
regulatory element activity (Figure 14 (A)) and are active in a range of
plants, and
CA 02507563 2005-05-13
direct the constitutive expression of a coding region of interest throughout a
plant
(Figure 14,(B)).
[0081 ] Therefore, the present invention alsb provides ~~for a chimeric
nucleic acid
construct comprising a regulatory element in operative association with, a
coding '
region of interest, the regulatory element comprising nucleotides 1660-1992 of
SEQ
ID NO:1 (or SEQ ID N0:22), or a duplicate thereof. , ~ ' '
,, , ,
[0082] Another regulatory element of the present invention includes, but is
not limited
to, a post-transcriptional or translational enh~rlcer regulatory element
localized
~lPt\ PP O - C,,;h T f ~~ F ",-P 7 "" 1A r;aoQ~ '7'7n ~ ~ n i.rn. ~~. ,
~....n .. ~.., a 's Iguw. i, mu~m.(imuW 20UZ-2GG-T of Slug IL lVll.l or GG, or
I
p,,~,wlapti,~iac 1_1 ~jø~ ~f ~F,Q TD N(]:21 a1~:3 r~ierr~d tn a3 "~T"_ The .t-
,,~5tvru~':wripiiv nui
or translational enhances regulatory element may also camprise the nucleotide
sequence as defined by nucleotides 1-141 of SEQ ID N0:2 (nucleotides 2084-2224
of ,
SEQ ID NO:1 or 22) or an analog thereof, or the element may comprise 70%
similarity (sequence identity) to the nucleotide sequence of nucleotides 1-141
of SEQ
ID N0:2 (nucleotides 2084-2224 of SEQ ID NO:1 or 22). This regulatory' element
'
also exhibits substantial homology with members of the RENT family of
repetitive
elements (see Figure 13 (C); nucleotides 495-635 or nucleotides 2084-2224 of
tCUP). ,
[0083] A shortened fragment of the NdeI - SmaI fragment, referred to as 0N,
dN,
deltaN, or tCUP delta, is also characterized within the present invention. ON
was
prepared by mutagenesis replacing the out of frame ATG (located at nucleotides
2087-2089, SEQ ID NO:1) within the NdeI-SmaI fragment (see Figure 10). ON
constructs with (SEQ ID N0:3) or without (SEQ ID N0:4) a Kozak consensus
sequence was also characterized (Tables 10, and 12) and found to exhibit
enhances
activity. Therefore, other cryptic regulatory elements of the present
invention include,
but are not limited to, post- transcriptional or translational enhances
regulatory
elements localized at nucleotides 1-97 of SEQ ID NO's:3 and nucleotides 1-86
of
SEQ ID NO's: 3 or 4. These post-transcriptional or translational enhances
regulatory
elements may comprise the nucleotide sequence as defined by nucleotides 1-86
of
31
CA 02507563 2005-05-13
SEQ ID NO's:3 or 4 (nucleotides 2091-2170 of SEQ ID NO:1) or an analog
thereof,
or the element may comprise 70% similarity to the nucleotide sequence of
nucleotides
1-86 of SEQ ID NO's:3 or 4 (nucleotides 2091-2170 of ~SEQ ID NO:1).
Furthermore,
these regulatory elements may comprise the nucleotide sequence as defined by
nucleotides 1-97 of SEQ ID N0:3 and comprising a Kozack sequence or an analog
thereof, or the element may comprise 70% similarity to the nucleotide sequence
of
nucleotides 1-97 of SEQ ID N0:3.
[0084] Furthermore, other regulatory elements of the present invention include
negative regulatory elements (for example located within an XbaI-BstYI
fragment as'
defined by Figure 5 (C); nucleotides 1-1660 of SEQ'ID NO:1), a transcriptional
enhancer_ localized within the BstYI-DraI fragment of Figure 5 (C)
(nucleotides 1660-
1875 of SEQ ID NO: i), a core promoter element located within the Dral-NdeI
fragment of Figure 5 (C) (nucleotides 1875-2084 of SEQ ID NO:1 or 22), a
transcriptional enhancer within the Dral to -62 fragment (nucleotides 1875-
19,92 of ,
aLQ ii iw~: l UI 22; i figures 5 (v) to (u)), or a regulatory element or post-
transcriptional element downstream of the transcriptional start site, for
example but
not limited to the NdeI-SmaI fragment (nucleotides 1-188 of SEQ ID N02) and
derivatives and fragments thereof (for example nucleotides 1-141 of SEQ ID
N0:2);
including ON (nucleotides 1-129 or 1-97 of SEQ ID N0:3, ONM (nucleotides 1-119
or
1-86 SEQ ID N0:4), and nucleotides 1-86 of SEQ ID N0:3 or 4 (nucleotides 2084
to
2170 of SEQ ID NO: l ). '
[0085] The following non-limiting list of fragments of SEQ ID NO:1 or 22 have
been
characterized and their utility demonstrated herein, nucleotides:
1660-1992 ("-394" to "-62" fragment) enhances expression of the -46 minimal
promoter of 35S, and a fragment of T1275 (see Bstl-GUS; Bstl-355, Bst2-GUS,
Bst2-35S, of Figure 5D);
1660-1875 (BstYI-DraI fragment; see Figure SC; and Table 6; -394 GUS-nos)
exhibits enhancer activity;
32
CA 02507563 2005-05-13
' '
1660-2224 (BstYI-SmaI fragment; see Figure SC; and Tables 5 and 6; -394-
GUS-nos) .also exhibits enhancer activity;
1724-2224 (Figures 13C, and Figure 14A, "tCUP RENT") exhibits regulatory
element activity and comprises .several regulatory elements (core promoter
element
and an translational enhancer element). Nucleic acid sequences that hybridize
to ~ ~ '
nucleotides 1724-2224 under stringent hybridization conditions and that
exhibit one or ' '
more than one regulatory element activity, or nucleic acid sequences that
exhibit , ' ~ , , ,, .
greater than 75% sequence identity with nucle,otides,'1724-2224 and that
exhibit one ,
Ui inorc than one regulatory element activities, arelmembers of the RENT
family
(SEQ ID NO's:21 and 22); ,
' ' ~ '
1875-2084 (DraI-NdpI fragment; core promoter element), see Figure SC and
Table 6 (-197-GUS-nosy; ~ ~ ,
1875-1992 (DraI - "-62" fragment) This fragment is shown to enhance
expression of the -46 minimal promoter of ~ 5 S, arid a fragment of T1275, as
shown in
~~
Figures SD (see Dral-GUS; Dra2-GUS; Dral-35S; Dra2-35S), and Figures SE-G
(Dral-355; Drat-35S), and functions as a transcriptional enhancer;
2084-2224 (NdeI-SmaI fragment, or "N"; Tables 10-12, Figure SB (+~0-GUS-
nos), Figure 7 (T127~-GUS-nos; 355-GUS-nosy, and Figure 11 (35S+N-GUS-nos)
exhibits translational regulatory element activity; and
2091-2170 (ON; see Tables 10-12) exhibits translational enhancer activity.
[0086] Therefore, the present invention is directed to an isolated nucleic
acid
sequence comprising a regulatory element selected from the group consisting of
a
nucleotide sequence:
defined by nucleotides I-1660 of SEQ ID NO:1 or 22 (XbaI-BstYI),
defined by nucleotides 1660-1992 of SEQ ID NO:1 or 22 (BstYI to -62),
JJ
CA 02507563 2005-05-13
defined by nucleotides 1660-1875 of SEQ ID NO:1 or 22 (BstYI-DraI),
defined by nucleotides 1660-2224 of SEQ ID NQ:1 or 22 (BstYI-SrnaI),
defined by nucleotides 1724-2224 of SEQ ID NO:1 or 22 (RENT),
defined by nucleotides 1875-2084 of SEQ ID NO:1 or 22 (DraI-NdeI),
I defined by nucleotides 1875-1992 of SEQ ID NO:1 or 22(Dral to -62),
. defined by nucleotides 2084-2224 of SEQ ID NO:1 or 22 (NdeI-Smal),
defined by nucleotides 2091- 2170 of SEQ ID NO:1 or 22 (N),
defined by nucleotides 1992-2042 of SEQ ID NO:1 or 22 (-62 to -12),
defined by nucleotides 415-2224 of SEQ ID NO:l or 22 (SphI- Smal),
defined by nucleotides 1040-2224 of SEn ID NOv 1 or 72 ~PctT- .(',s,.yl h gird
'
defined by nucleotides 1370-2224 of SEQ ID NO:1 or 22 (SspI-Smal).
[0087] The present invention also provides an isolated nucleic acid sequence
comprising a regulatory element selected from the group consisting of a
nucleotide
sequence:
that hybridizes to nucleotides 1-1660 of SEQ ID NO:1 or 22 or a compliment
thereof,
that hybridizes to nucleotides 1660-1992 of S$Q ID NO:1 or 22_ or a
compliment thereof,
that hybridizes to nucleotides 1660-1875 of SEQ ID NO:I or 22 or a
compliment thereof,
34
CA 02507563 2005-05-13 .
that hybridizes to nucleotides 1724-2224 of SEQ ID NO:1 or 22 or a
compliment thereof, ' . . ~ , ,
thathybridizes to nucleotides 1875-2084 of SEQ ID NO:1 or 22 or a
compliment thereof,
that hybridizes to nucleotides 1875-2224 of SEQ ID NO:1 or 22 or a
compliment thereof,
that hybridizes to nucleotides 1875-1992 of SEQ ID NO:I or 22 or a
I compliment thereof, ~ ' ,
that hybridizes to nucleotides 2084-2224 of SEQ ID NO:1 or 22 nr a
compliment thereof, and ,
that hybridizes to nucleotides 2091-2170 of SEQ ID N0:1 or 22 or a
compliment thereof, '
wherein hybridization is under a co_n_di_tion set_ectPd from the grrnop
rnngigting of:
hybridizing overnight (16-20 hours) in a solution comprising 7% SDS, O.SM
NaP04 buffer at pH 7.2, and 10 mM EDTA at 65°C and washing for one hour
at 60°C
in a solution comprising 0.1 X SSC and 0.1% SDS;
hybridizing overnight (16-20 hours) in a solution comprising ?% SDS, O.SM
NaP04 buffer at pH 7.2, and 10 mM EDTA at 65°C and washing for one hour
at 65°C
in a solution comprising 2 X SSC and 0.1% SDS; and
hybridizing overnight (16-20 hours) in a solution comprising 4 X SSC at
65°C
and washing one hour in 0.1 X SSC at 65°C, and
wherein the regulatory element exhibits regulatory element activity and is
capable of
mediating the transcriptional or translational efficiency of a transcript
encoding a
coding region of interest that is operatively linked thereto.
3~
CA 02507563 2005-05-13
[0088] Furthermore, the present invention
provides an isolated nucleotide sequence
comprising nucleotides defined by
the nucleotide sequence of SEQ ID
N0:22, or a
compliment thereof comprising the the positions
following nucleotides at indicated
in
'
,
Table 1 a. .
Table la: Identification of nucleotides ly and
of the RENT fami their
positions
, ~ '
within within SEQ ID N0:22 . . , ~ ' '.
Position* Nucleotide . '
~
~. . _ ,
' tCUP RENT1 RENT2 ~ RENT3 RENTS RENT?
1744 . C C ~ C ' ~' C , C A
1749 A A A ' T' A A
1?50 T T T . T T C
1751 C T T~ T T ~ T
1763 G A A ~ A A A
1764 C T T T T T
1767 A T A A A A
1783 T T C T T T
1788 T A T T T C
1789 A A A T A A
1790 C G C C C C
1794 C T C C C C
36
CA 02507563 2005-05-13
1799 T T C ~T T T
2000 G T G ~ ~, G, G , ~ A
~
' 1807 G G A G G G
' 1811 T T T T T C
1812 T C 'T T C C
1823 , T T T ~ ~~T G ~ T
1 O'1 ~ T T
1 O.C.,'t 1 1 j ~ j
1827 T - T ~T T T
1828 A A G A A A
1834 T A T T T ~ T
1835 A C A A A A
1839 G A G G G G
1842 A G A A A A
1843 G A G G G , G
1847 A G A A A A
1851 C T C C C C
1863 T T T C T C
1866-7 - C C C C A
1873 T C T T T T
37
CA 02507563 2005-05-13
~,
s
1883 T A T ~ T T ~ T
1886 G G G ~ . , A G G ,
,
a ,
1897 G G ~ G G G A ' '
.
1897-8 - C C -C C A ~ '
' '
1901 C ,, , ~ ,-
- C , , . - - ,,
1903 A A A A A . '
~ ~G
,
i 907 G A ~A G . . A A
~ ~
~. ,
1912 A ~ T -
A A -
1913 A A 'A - A ~ - '
1914 T T T , ' - T
.
1915 T T T - T -
1916 T C T - T -
1917 C C C - C -
1918 A A A ~ - A -
1919 A A A - A -
1920 A A A - A -
1921 T T T - T -
1922 T T T - T -
1923 C C C - C -
38
CA 02507563 2005-05-13 '
1927 ' C ~ T ~C C C '
T
. , ,
1930 T G G v C ~ G ~ G ,
1931 C C A ~ C C C
1932 C C C ' C I T C ,
1934 G G ~ G T G G
'
,
' ~ w1939 , ~ A A ' ~A G ' A
A ,
1947-8 - A ~ A ~A A ,
A
1949 T T T ~T T C
1950 A G A A A A
.. , ,
' 1952 C C C T C C
1954 A ~= G v T
1956 C T C C C C
GCTTTTC TCTTATC GCTTATC GCTTATC GCTTATC
1964-5 -
CAACCC CAACCC CAACCC CAACCC .CAACCC
1966 G G A G G G
1969 G G G G T G
1970 G G A G G A
1973 T T T C T T
1976 C C C - C T
39
CA 02507563 2005-05-13
1990 C C T C C ,C
1991 ~ C T T ~ , C , C C '
2012 C G , A A , A ~ G
2026 T T C ~ T II T T
,, ~ .
2029 T C ', C , , . _ C '
C y ~,
,,
2031 ~ C _ T , ~ ' C . ,
,
2032 T _ , G T T T '
2038 A A A A A C
2051-2 - C C G C ~ C '
2054 T C C _ _ _ .
.
?n5~ ~ r~ ~ A C A
2058 T C C C C C ,
2059 C T C C C C
2066 A G A ~ A A A
2086 A C C C C C
2089 G - - A - A
2090 A - - T - T
2091 A A A _ A _
2092 T T T - T _
CA 02507563 2005-05-13
' ~ ~ , S
2171 A C C C C ~C '
2172 ~ A C C ~. , C , C C . ' ,
a " ,
2173 T A ~ A A ' A A '
,, , ,
2174 A C C CI~ C C ~ ~ '
,, , ~ ,
2181 T C ', C , , .C C C
2185 ~ C A A , ~A ~ A ' A
~
,,
,.
2186 A G G G G ,
' G
2189 C - ~' ' G - G
-
2190 C - - A
2191 G - ~- T - T
2192 T - - C
2193 G G - - G - ,
2194 G A G - A -
2195 A T A - T -
2196 A C - - C _
2197 A C - C C C
2198 C C - C C C
2199 C C - C C C
2200 T G - G G G
41
CA 02507563 2005-05-13
2201 T G ~ - ~G G G
2202 A G - G , G ' G
2204 A G - G G G
2205 C G - G ~G G ,
2209 C G - G I G G
2210 C T - ~, T T T
' ,
2211 T C T C C C
2214 C T C ~T T T
2215 T T C T T T
2216 T A G A A ~ A
2617 T T - 1 L L
2218 G G - G G G
2219 C - - T - T
2220 T - - N - . T
2221 C - - A - A
2222 T T - - T -
2223 T T - - T -
2224 A A - - A -
* position within SEQ ID N0:22
42
CA 02507563 2005-05-13
wherein the nucleotide sequence exhibits regulatory element activity and is
capable of
conferring ,or enhancing expression on a coding region of interest linked in
operative ,
association therewith. '
[0089] An "analogue" of the above identified regulatory elements includes any
~ '
substitution, deletion, or additions to the sequence of a regulatory element
provided ' ' ,
that said analogue maintains at least one regulat'or'y property associated
with the ~ ' ~. , ,,
activity of the regulatory element. Such properties include directing organ
specificity, '
tissue specificity, or a combination thereof, pr.temporal activity, or
developmental .
activity, or a combination thereof, or other regulatory attributes including,
negative
regulatory elements, enhancer sequences, or sequences that affect stability of
the '
transcription or translation complexes or stability of tl~e transcript.
[0090] ~he present invention is further directed to a chimeric gene construct
containing a DNA of interest operatively linked to,the regulatory element of
the
present invention. Any exogenous gene can be used and manipulated accdrding to
th'e
present invention to result in the expression of said exogenous gene. A DNA or
coding region of interest may include, but is not limited to, a gene encoding
a protein, ,
a DNA that is transcribed to produce antisense RNA, or a transcript product
that
functions in some manner that mediates the expression of other DNAs, for
example
that results in the co-suppression of other DNAs ,or the like. A coding region
of
interest may also include, but is not limited to, a gene that encodes a
pharmaceutically
active protein, for example growth factors, growth regulators, antibodies,
antigens,
their derivatives useful for immunization or vaccination and the like. Such
proteins
include, but are not limited to, interleukins, insulin, G-CSF, GM-CSF, hPG-
CSF, M-
CSF or combinations thereof, interferons, for example, interferon-a,
interferon-l3,
interferon-i, blood clotting factors, for example, Factor VIII, Factor IX, or
tPA or
combinations thereof. A coding region of interest may also encode an
industrial
enzyme, protein supplement, nutraceutical, or a value-added product for feed,
food, or
both feed and food use. Examples of such proteins include, but are not limited
to
43
CA 02507563 2005-05-13
proteases, oxidases, phytases, chitinases, invertases;,lipases, cellulases,
xylanases,
enzymes involved in oil biosynthesis etc.
[0091 ] The ~chimeric gene construct of the present invention can further
comprise a 3'
untranslated region. A 3' unfranslated region refers to that portion of a gene
comprising a DNA segment that contains a polyadenylation signal and any other
regulatory signals capable of effecting mRNA processing or gene expression.
The
,.
polyadenylation signal is usually characterized by effecting the addition of
polyadenylic acid tracks to the 3' end of the mRNA'precursor. Polyadenylation
. ' '
signals are commonly recognized by the presence of homology to the canonical
form' .
5' AATAAA-3' although variations are not uncommon.
[0092] Exaanples of suitable 3' regions are the 3' transcribed non-translated
regions
containing a polyadenylation signal of Agrobacterium tumor inducing (Ti)
plasmid
genes, such as tr~e nopaline synthase (Nos gene) and plant genes such as the
soybean
y , ,
storage protein genes arid the small subunit of the ribulose-1, 5-bisphosphate
carboxylase (ssRUBISCO) gene. The 3' untranslated region from the structural
gene
of the present construct can therefore be used to construct chimeric genes for
' expression in plants.
[0093] The chimeric gene construct of the present invention can also include
further
enhancers, either~translation or transcription enhancers, as may be required.
These
enhancer regions are well known to persons skilled in the art, and can include
the
ATG initiation codon and adjacent sequences. The initiation codon must be in
phase
with the reading frame of the coding sequence to ensure translation of the
entire
sequence. The translation control signals and initiation codons can be from a
variety
of origins, both natural and synthetic. Translational initiation regions may
be
provided from the source of the transcriptional initiation region, or from the
structural
gene. The sequence can also be derived from the regulatory element selected to
express the gene, and can be specifically modified so as to increase
translation of the
mRNA.
44
CA 02507563 2005-05-13
~, , , s
[0094] To 'aid in identification of transformed plant cells, the constructs of
this
,
invention may be further manipulated. to include plant selectable markers.
Useful .
selectable markers include enzymes which provide for resistance to an
antibiotic such
" ,
. ,
as gentamycin, hygromycin, kanamycin, and tlie, like. Similarly, enzymes
providing '
for production of a compound identifiable.by colour c~iange such as GUS ~ 1 ,
. '
(13-glucuronidase), or luminescence, such as luciferase are useful.
' ' '' ~ ' .
[0095] Also considered part of this invention are'transgenic plants, trees,
yeast, ~ ' ~:
bacteria, fungi, insect and animal cells. containing tl~t chim~ric gene
construct
comprising a regulatory element of the presyt invention. 'However, it is to be
'
understoad that the regulatory elements of the present invention may also be
combined with coding region of interest for expression within a rar_ge of host
'
organisms that are amenable to transformation. Such,organisms include, but are
not
limited to: ' ,
~ plants, both monocots and dicots, for example, corn, cereal plants, wheat,
barley, oat, tobacco, Brassica, soybean, pea, alfalfa, potato, ginseng,
Arabfdopsis;
~ trees, gymnosperms and angiosperms, including both hardwood and softwood
trees, for example peach, plum, spruce;
~ yeast, fungi, insects, animal and bacteria cells.
[0096] Methods for the transformation and regeneration of these organisms are
established in the art and known to one of skill in the art and the method of
obtaining
transformed and regenerated plants is not critical to this invention.
[0097] In general, transformed plant cells are cultured in an appropriate
medium,
which may contain selective agents such as antibiotics, where selectable
markers are
used to facilitate identification of transformed plant cells. Once callus
forms, shoot
formation can be encouraged by employing the appropriate plant hormones in
accordance with known methods and the shoots transferred to rooting medium for
CA 02507563 2005-05-13
s
regeneration of plants. The plants may then be used to establish repetitive
generation, either from seeds or using vegetative propagation techniques.
[0098] The constructs of the present invention can be introduced
into'plant~cells using
Ti plasmids, Ri plasmids, plant virus,vectors, direct DNA transformation,
micro=
injection, electroporation, etc. For reviews of such techniques see for
example
Weissbach and Weissbach, Methods for Plant Molecular Biology, Academy Press,
New York VIII, pp. 421-463 (1988); Geierson and Corey, Plant Molecular
Biology, , ~ ~; , ,,
2d Ed. (1988); and Miki and Iyer, Fundamentals of Qene Transfer in Plants. In
Plant
Metabolism, 2d Ed. DT. Dennis, DH Turpin,'DD Lefebrve; DB Layzell (eds),
Addison
Wesly, Langmans Ltd. London, pp. 561-579 (197). The present invention further
includes'a suitable vector comprising t'he chimeric gene canstruct. '
[0099) When specific sequences are referred to in the present invention, it is
understood that these sequences include within their scope sequences that are
'°substantially homologous" to the specific sequences, or sequences or
a compliment ,
of the sequences hybridise to one or more than one nucleotide sequence as
'defined ~ '
herein under stringent hybridisation conditions. Sequences are "substantially
homologous" when at least about 70%, or more preferably 75% of the nucleotides
match over a defined length of the nucleotide sequence providing that such
homologous sequences exhibit one or more than one regulatory element activity
as
disclosed herein. For example which is not to be, considered limiting, the
RENT
family of nucleotide sequences as defined herein exhibits greater than about
75%
sequence similarity with a fragment (nucleotides 1724 to 2224) of the
nucleotide
sequence of SEQ ID NO:1 or 22. Furthermore, members of the RENT family also
hybridise with the nucleotide sequence defined by SEQ ID NO: l or 22 under
stringent
hybridisation conditions and exhibits one or more than one regulatory element
activity.
[00100] Such a sequence similarity may be determined using a nucleotide
sequence comparison program, such as that provided within DNASIS (using, for
46
CA 02507563 2005-05-13
example but not limited to, the following parameters: GAP penalty 5, #of top
diagonals 5, fixed GAP penalty 10, k-tuple 2, floating gap 10, and window size
5),.
However, Qther methods of alignment of sequences'for comparison are weld-known
in
' the art for example the algorithms of Smith & Waterman (Adv. Appl. Math.
2:482,
1981), Needleman & Wunsch (J. Mol. Biol. 48:443, 1970), Pearspn & Lipman
(Proc.
Nat'l. Acad. Sci. USA 85:2444, 1988), and by computerized implementations of
these
' ,
algorithms (GAP, BESTFIT, FASTA; and BLAST, available through the NIH.), or by
, ,
manual alignment and visual inspection (see, e.g., Current Protocols in
Molecular
Biology, Ausubel et al., eds. 1995 supplement), or using Southern or Northern
,,
~y~r~d'zatio:~ uirder Strlngcnt C,onditlyrlj (see i~ianiatis ei ai., in
i~oiecuiar Cloning (A '
~. ".~
Laboratory Manual), Cold Spring Harbor Laboratory, 1982) to the nucleotide
sequence of SEQ ID NO's:l, 5, 6, 7, 8, 9, 21 or 22 provided that the sequences
maintain at least one regulatory property or regulatory element activity, as
defined
herein. Preferably, sequences that are substantially homologous exhibit at
least about
80% and most preferably at least about 90% sequence similarity over a defined
length
of the molecule. ,
[001 O 1 ] The DNA sequences of the present invention thus include the DNA
sequences of SEQ ID NO's:l, 5, 6, 7, 8, 9 21 or 22, their regulatory regions
and
fragments thereof, as well as analogues of, or nucleic acid sequences
comprising
about 70% similarity with the nucleic acids, or fragments thereof, as defined
in SEQ
ID NO:1, 5 to 9, 21 and 22. Sequences that are "substantially homologous"
include
any substitution, deletion, or addition within the sequence.
[00102] An example of one such stringent hybridization conditions may be
overnight (from about 16-20 hours) hybridization in 4 X SSC at 65°C,
followed by
washing in 0.1 X SSC at 65°C for an hour, or 2 washes in 0.1 X SSC at
65°C each for
20 or 30 minutes. Alternatively an exemplary stringent hybridization condition
could
be overnight (16-20 hours) in 50% formamide, 4 X SSC at 42°C, followed
by washing
in 0.1 X SSC at 65°C for an hour, or 2 washes in 0.1 X SSC at
65°C each for 20 or 30
minutes, or overnight (16-20 hours), or hybridization in Church aqueous
phosphate
47
CA 02507563 2005-05-13
buffer (7% SDS; O.SM NaP04 buffer pH 7.2; 10 mM EDTA) at 65°C, with 2
washes
either at 50°C in 0.1 X SSC, 0.1% SDS for 20 or 30 minutes each, or 2
washes at
65°C in 2 X SSC, 0.1% SDS for 20 or 30 minutes each for unique seduence
regions.
[00103] Analogues also include those DNA sequences which hybridize, to the
sequence of SEQ ID NO:1, 5, 6, 7, 8, 9, 21 or 22 or a fragment thereof, under
relaxed ~ , '
hybridization conditions, provided that said sequences maintain at least one
regulatory
property of the activity of the regulatory element. ' Examples of such relaxed
' ' ' , ,
hybridization conditions includes overnight (16-20 fours) hybridization in 4 X
SSC at
. .
50°C, with 30-40% formamide at 42°C, or 65°C in 2 X SSC,
0.1% SDS for example
for analysis of repetitive regions as described hererin
[00104] The specific sequences, referred to in the present invention, also
include sequences which are "functionally equivalent" to said specific
sequences. ~ In
the present invention functionally equivalent sequences refer to sequences
which
although not identical to the specific sequences provide the same or
substantially the
same function. DNA sequences that are functionally equivalent include ariy
substitution, deletion or addition within the sequence. With reference to the
present
invention functionally equivalent sequences will preferably direct the
expression of an
exogenous gene constitutively.
[00105] The results presented in the examples indicate that the constitutive
expression of GUS activity in the plant T1275 is regulated by a cryptic
regulatory
element. Similarly, other experiments indicate that homologs of the cryptic
regulatory
element (for example members of the RENT family) are also effective in
obtaining
constitutive expression of a coding region of interest under their control.
RNase
protection assays performed on the region spanning the regulatory element and
downstream region did not reveal a transcript for the sense strand (see Figure
8, Table
2). RNase protection assays were performed using RNA from organs of
untransformed tobacco and probes that spanned the T1275 sequence from about -
2055
by to +1200 by relative to the transcriptional start site. In all tissues
tested (leaf, stem,
48
CA 02507563 2005-05-13
root, flower bud, petal, ovary and developing seed) protected fragments were
not
detected, in the sense orientation relative to the GUS coding region,
,with~all probe
(Figure 8), and indicates that the site was the same in each organ.
Furthermore,
GenBank searches revealed no significant sequence similarity with the T127S
sequence. An amino acid identity of about 66% with two open reading frames on
the
antisense strand of the genomic sequence of T127S (between about -1418 and -
1308;
nucleotides 636-746 of SEQ ID N0:1; and between about -541 and -395;
nucleotides
1513-1659 of SEQ ID NO:1 relative to the transcriptional start) and an open
reading
frame of a partial Arabidopsis expressed sequence (GenBank Accession No.
W43439)
;~°as ldWtiiied. T he sequence which lies downstream of sequences at
tile T-DNA ~ I
insertion point in untrar~sforined tobacco shows no significant similarity,in
GenBank
searches. These data suggest that this region is silent in untransformed
plants and that
the insertion of the T-DNA activated a cryptic regulatory element.
[00106] Similar RNase protection assays using probes from tCUP (T127,S)
against members of the RENT family of sequences (SEQ ID NO's: S to ~9)
indicates
that thcsc SequenGe$ are dl5u SllCni lri untransIOrmeCl plants
[00107] Southern analysis indicates that the 2.2 kb regulatory region of T1275
does not hybridize with DNA isolated from soybean, potato, sunflower;
Arabidopsis,
B. napus, B. oleracea, corn, wheat or black spruce. However, transient assays
indicate
that this regulatory region can direct expression of the GUS coding region in
all plant
species tested including canola, tobacco, soybean, alfalfa, potato, Ginseng,
peach, pea,
Arabidopsis, B. napus, white spruce, corn, wheat, oat and barley (Table 3),
indicating
that this regulatory element is useful for directing gene expression in both
dicot and
monocot plants. A fragment of the T1275 (tCUP) regulatory region that exhibits
substantial homology with a segment of the RENT family of repetitive elements,
and
the corresponding fragments from the RENT nucleotide sequences, for example,
but
not limited to SEQ ID NO's: S to 9, and 21 are also active in other species,
for
example but not limited to pea and Arabidopsis (see Figure 14).
49
CA 02507563 2005-05-13
[00108] The following fragments of the members of the RENT family (see SEQ
ID N0:21), and there corresponding fragments of SEQ ID NO:l, have been ,
characterized, and their utility demonstrated in thepresent ,invention. For
example,
the fragment comprising nucleotides from SEQ' ID NO:l or 22 of:
.
1724-2224 and nucleotide sequences that are characterized as having greater ~
~ .
than 75% sequence identify with nucleotides 1724-2224 of SEQ ID NO~1 or 22
(see ' '.
Figure 13C) exhibit regulatory element activity (e:g. Figure 14(A)); , ~ ' ';
, ,,
1875-2086 (DraI-NdeI fragment; core promoter element), see Figure 5C and
. ,
Table 6 (-197-GUS-nosy;
1875-1992 (DraI -62 fragment) - this fragment enhances expression of the -46
minimal promoter of 355, and a fragment of T1275, as shown in Figures 5D (see,
Dral-GUS; Dra2-GUS; Dral-355; Dra2-35S), and Figures 5E-G (Drat-355; Dra2-
35S), and functions as a transcriptional enhancer;
2084-2224 (NdeI-SmaI fragment; or "N"; Tables 10-12, Figure 5R r+30_GTT~~
nosy, Figure 7 (T1275-GUS-nos; 35S-GUS-nosy, and Figure 11 (35S+N-GUS-nos) a
translational enhancer; and
2091-2170 (~N) see Tables 10-12; a translational enhancer.
[00109] The transcriptional start site of T1'275 (tCUP) was delimited by RNase
protection assay to a single position about 220 by upstream of the
translational
initiation codon of the GUS coding region in the T-DNA. The sequence around
the
transcriptional start site exhibits similarity with sequences favored at the
transcriptional start site compiled from available dicot plant genes (T/A T/C
A+1 A
C/A C/A A/ClT A A A/T). Sequence similarity is not detected about 30 by
upstream
of the transcriptional start site with the TATA-box consensus compiled from
available
dicot plant genes (C T A T A A/T A T/A A).
CA 02507563 2005-05-13
[00110] ~ Deletions in the upstream region indicate that negative regulatory
elements and enhancer sequences exist within the frill length regulatory
region. FQr
example deletion of the 5' region to BstYI (-394 relative to the
transcriptional start
site; position 1660 of SEQ ID NO:1 or 22) resulted in a 3 to 8 fold increase
in
expression of the gene associated therewith (see Table 6 in Examples, and
Figure 5
(C)), indicating the occurrence of at least one negative regulatory element
within the
XbaI-BstYI portion of the full length regulatory element. Other negative
regulatory
elements also exist within the XbaI- BstYI fragment of T1275 as removal of
an~XbaI-
~Sii iagWV:ut aiw reSUitcd in iuCreascd actiYity (-i304-GUS-nos; Table 6,
Examples,
and Figure 5, comprising a deletion of nucleotides 1-1040 of SEQ ID NO:1 or
22).~ ,
[00111 ] An enhancer is also localized within the BstYI-DraI fragment of tCUP
as removal of this region results in a 4 fold loss in activity of the
remaining regulatory
region (-197-GUS-nos; Table 6, Examples, and Figure 5, comprising a deletion
of
nucleotides 1-1875 of SEQ ID NO:1 or 22). In addition to the -197 to -62
region ,
(corresponding to nucleotides 1875 to 1992 of SEQ ID NO:1 ) exhibiting
enhancer-
like properties, the region spanning -394 to -62 (corresponding to nucleotides
1660 to
1992 of SEQ ID NO:1) also exhibit similar prope ~ies. When the -19 7 to -62
(nucleotides 1875-1992 of SEQ ID NO:1 or 22) and -394 to -62 (nucleotides 1660-
1992 of SEQ ID NO:1 or 22) fragments of T1275 canstruct are fused with the -46
minimal promoter of 35S, the promoter activities were enhanced to about 150
fold
(Dral-355 Figure 5 (D)). Duplication of the -197 to -62 (Dra2-GUS; Figure 5
(D)), or
the -394 to -197 (labelled as Bstl in Figure 5 (D)) fragments, or a
combination of
these two fragments, resulted increased regulatory element activity when
placed in
association with a regulatory element fragment, for example, T1275 (Bst2-GUS;
Figure 5 (D)) or 355 (Bst2-35S).
[00112] 5' deletions of the regulatory element (see Figures 5(A) and (B) and
analysis by transient expression using biolistics showed that the regulatory
element
was active within a fragment 62 by from the transcriptional start site
indicating that
the core promoter has a basal level of expression (see Table 5, Examples; and
Figures
51
CA 02507563 2005-05-13
' "
s
(H) and (I)). Deletion of a fragment containing the transcriptional start site
(see -
,
62(-tsr)-GLJS-nos in Figures 5 (B), (H) and (I); Table 5, Examples) reduced ,
expression dramatically in transgenic tomato, however deletions to +30 did
eliminate
" ,
expression indicating that the region defined fr6m about -12 to about +30 by '
contained the core promoter. Deletion of sequences surrounding the
trasncriptional ,
start site, reduced activity to about 2% of the activity associated with the -
62-GUS ~ '
construct, indicating that the trasncriptional start site sequence is required
for tCUP
,,.
regulatory element activity. DNA sequence searches did not reveal conventional
core ' ~ ''
. . ~ , ,
prp_rn__YtPr mntifc fpynd in plant ~r.ene5 ~u~h a~ i~~W T, ETA bur. ' , ~ , .
" ,
" ~ . .
[00113] Substitutian of nucleotides at'-30 to -24, of -62-GUS-nos, with the
TATA-box sequence TATATAA (~igures 5 (D)~and (H),"increased care promoter
acti wiry abo~zt 3 fold (r figure 5 (1). Addition of,a GCC-box sequence (Hart
et ai.,1,993;
Ohme-Takagi and Shinshi, 1995) to -62-GUS-nos resulted in about a four fold
increase in activity (see Figure 5 (I)). The results presented in Figures 5
(D) and (I)
demonstrate that the regulatory elements of the present invention may be
modulated '
through a variety of modifications including duplication of fragments that
exhibit
enhar~cer or silencer activity, or by substituting, inserting, or adding
regulatory
elements to enhance or silence tCUP regulatory element activity.
[00114] A number of the 5' regulatory element deletion clones (Figure 5 (C))
were transferred into tobacco by Agrobacterium-mediated transformation using
the
vector pRD400. Analysis of GUS specific activity in leaves of transgenic
plants (see
Table 6, Examples) confirmed the transient expression data down to the -197
fragment (nucleotides 1857-2224 of SEQ ID NO:1). Histochemical analysis of
tobacco organs sampled from the transgenic plants indicated GUS expression in
leaf,
seeds and flowers. Histochemical analysis of Arabidopsis organs revealed GUS
activity in leaf, stem flowers and silques when the promoter was deleted to
the -394
and -197 fragments (see Figures 5 (E) to (G)).
52
CA 02507563 2005-05-13
[0011 S] As indicated above, a fragment of the regulatory element tCUP
(T1275) exhibits substantial homology with a large family of repetitive
elements ,
within N. ta.bacum. These homologous sequences (SEQ~ ID NO's: 5 to 9; RENT 1,
2,
3, 5 and 7) also exhibit regulatory activity as determined by an increase in
the
expression of GUS in pea protoplast assays (Figure ~14 (A)). This,region (-394
tCUP-
GUS) was also found to drive the constitutive expression of a coding region of
interest
in transgenic Arabidopsis (Figure 14 (B)). Therefore, the present invention
also
describes the regulatory elements associated with members of the RENT family
of
repetitive elements including tCUP (T1275). The consensus sequence for
_m__e_m__bP_rs ~f
,,
~P pFNT f~rrily i5 prOVlded in Fig'akes i3(C) aad i~3(D).
[00116] Expression of GUS, under the contral of T1275 or a fragment thereof,
or the modulation of GUS expression arising from x'1275 or a fragment thereof,
has
been observed in a range of species including corn, wheat, barley, oat;
tobacco, ,
Brassica, soybean, alfalfa, pea, potato, Ginseng, Arabidopsis, peach, spruce,
yeast, ,
fungi, insects and bacterial cells ('Table 3, Examples, and Figures 14 (A),
and (~)).
Occurrence of a post-transcriptional regulatory element in the T1275
nucleotide
sequence
[00117] A comparison of GUS specific activities in the leaves ef transgenic
tobacco SR1 transformed with the T1275-GUS-nos gene and the 35S-GUS-nos genes
revealed a similar range of values (Figure. 6(A)). Furthermore, the GUS
protein
levels detected by Western blotting were similar between plants transformed
with
either gene when the GUS specific activities were similar (Figure. 6(C)).
Analysis of
GUS mRNA levels by RNase protection however revealed that the levels of mRNA
were about 60 fold (mean of 13 measurements) lower in plants transformed with
the
T1275-GUS-nos gene (Figure 6(B) suggesting the existence of a post-
transcriptional
regulatory element in the mRNA leader sequence.
[00118] Further analysis confirmed the presence of a regulatory sequence
within the NdeI-SmaI fragment of the mRNA leader sequence that had a
significant
53
CA 02507563 2005-05-13
impact on the level of GUS specific activity expressed in all organs tested.
Deletion
of the NdeI-SmaI fragment (nucleotides 2084-2224 bf SEQ ID N0:1 or 22) from
t$e
T1275-GUS-nos gene (Figure 7) resulted in about a'46-fold reduction in the
amount
of GUS specific activity that could be detected in leaves of transgenic
tobacco cv
Delgold (see Table 7). Similar~results were also observed in the t;ansgenic
tobacco
cultivar SRl and transgenic alfalfa (Table 7). Addition of the same fragment
to a
35S-GUS-nos gene construct (Figure 7) increased the amount of GUS specific
activity
by, about 5-fold in transgenic tobacco and a higher amount in transgenic
alfalfa '(see
Table 7, ). TnCreage4l GTJC u~t, ~',ty' :'Jas vbSCr ved iil urgalis of tobaccU
dIld alIalIa plants
tranformed with constructs containing NdeI-SmaI fragment (Table 8 and 9): This
data is cuiisisterit wit'n the presence of a post-transcriptional regulatory
element in this
fragment.
[00119) A modulation of GUS activity was noted in a variety of species that
were transformed with a regulatory element of the present invention. For
example but
not necessarily limited to; the NdeI-SmaI fragment of T1275 (also referred to
a~ "N")
and derivatives or analogues thereof, produced an increase in activity within
a variety
of organisms tested including a range of plants ( T ables 3 and 10, and Figure
11),
white spruce (a conifer; Table 11 ) and yeast (Table 12).
[00120] A shortened fragment of the NdeI-SmaI fragment, (referred to as
"0N", "dN", or "deltaN") was produced that lacks the out-of frame upstream ATG
at
nucleotides 2087-2089 of SEQ ID NO:l (see Figure 10(A) and (B)). Constructs
comprising T1275(~N)-GUS-nos yielded 5 fold greater levels of GUS activity in
leaves of transgenic tobacco compared to plants expressing T1275-GUS-nos.
Furthermore, in corn callus and yeast, ON significantly increased GUS
expression
driven by the 35 S promoter (Figure 119 and Table 10).
[00121 ] The Ndel Smal regulatory elements situated downstream of the
transcriptional start site functions both at a transcriptional, and post-
transcriptional
level. The levels of mRNA examined from transgenic tobacco plants transformed
54
CA 02507563 2005-05-13
with either T1275-GUS-nos, T1275-N-GUS-nos, 35S-GUS-nos, or 35S+N-GUS-nos,
are higher in transgenic plants comprising the Ndel Smal fragment under the
control
of the T1275 regulatory element but lower in those under control of the 35S
promoter,
than in plants comprising constructs that lack this region (Figures 9 (A) and
(B)). '
This indicates that this region functions by ether modulating transcriptional
rates, or
the stability of the transcript, or both. ,
[00122] The Ndel Smal region also functions posh-transcriptionally. The ratio
~ ' ,,
of GUS specific activity to relative RNA level,in indiwidual'transgenic
tobacco plants
that lack the Ndel Smal fragment is lower, arid when averaged indicates an
eight fold .
reduction in GUS activity per RNA, Char. in plants comorisir_g this rPgion_
lFigurP ~?
(C)). Sirizilarly, an increase, by an average of six fold, in 'GUS specific
activity is'
observed when the Ndel Smal region is added within xhe 35S untranslated
region,
(Figure 9 (C)). The GUS specific activ'ity:relative RNA levels are similar in
constructs 'containing the Ndel Smal fragment (tCUP-GUS-nos and 35S-~N-GT~TS-
nos). These results indicate that the Ndel Smal fragment (nucleotides 2084-
2224 of
SEQ ID NO:I or 22) modulates gene expression post-transcriptionally. Further
experiments suggest that this region is a novel translational enhancer.
Translation of
transcripts in vitro demonstrate an increase in translational efficiency of
RNA
containing the Ndel to Smal fragment (see Table 13). Furthermore, the levels
of
protein produced using mRNAs comprising the Ndel Smal fragment are greater
than
those produced using the known translational enhancer of Alfalfa Mosaic Virus
RNA4. These results indicate that this region functions post-
transcriptionally, as a
translational enhancer.
(00123] One or more of the constitutive regulatory elements described herein
may be used to drive the expression within all organs or tissues, or both of a
plant of a
coding region of interest, and such uses are well established in the
literature. For
example, fragments of specific elements within the 35S CaMV promoter have been
duplicated or combined with other regulatory element fragments to produce
chimeric
regulatory elements with desired properties (e.g. U.S. 5,491,288; US
5,424.200; US
CA 02507563 2005-05-13
' 5,322,938; US 5,196,525; US 5,164,316). As indicated above; the constitutive
' regulatory element or a fragment thereof, as defined'herein, may also be
used along
with other regulatory element, enhancer elements, or fragments thereof,
translational
' enhancer elements or fragments thereof in order to control gene expression.
Furthermore, oligonucleotides of 18 bps or longer are useful as prpbes, for
example to
identify other members of the RENT family of repetitive sequences, or as PCR '
' ,
primers in identifying or amplifying related DNA or RNA sequences in other
tissues , .
or prgamsms.
[00124] I Thus this invention is directed to a ccanstitutive regulatory
e1_ement~ .
associated regulatory elements identified within the tCUP nucleotide sequence
fSEO
ID NO:1 or 22), and combinations comprising one or more than one of these
regulatory elements. further this invention is directed to such regulatory
elements and
combinations thereof, in a cloning vector, wherein the coding region of
interest _is
under the control of the regulatory element and is capable of being expressed
its a ,
plant cell transformed with the vector. This invention further relates to
transformed
plant cells, transgenic plants regenerated from such plant cells, and seeds
produced
tom uiiese plarsts. The regulatory element, and regulatory element-gene
combination
of the present invention can be used to transform any plant cell for the
production of ,
any transgenic plant. The present invention is not limited to any plant
species.
[00125] Therefore, the regulatory elements of the present invention may be
used to control the expression of a coding region of interest within desired
host
expression system, for example, but not limited to:
~ plants, both monocots and dicots, for example, corn, tobacco, Brassica,
soybean, pea, alfalfa, potato, ginseng, wheat, oat, barley, Arabidopsis;
~ trees, for example peach, spruce;
~ yeast, fungi, insects, and bacteria.
56
CA 02507563 2005-05-13
.
,~ , ,
[00126] ~ Furthermore, the regulatory elements as described herein may be~
used .
in conjunction with other regulatory elements, such as 'tissue specific,
inducible or .
constitutive promoters, enhancers, or fragments thereof, and the like. For
example,
,~ ,
the regulatory region or a fragment thereof as defined herein maybe used to
regulate '
gene expression of a coding region of interest spatially"and
developmentally.within a ,
plant of interest or within a heterologous expression system, for example
yeast,
,
insects, or fungi expression systems. Regulatory~regions or fragments thereof,
~,~ ,
including enhancer fragments of the present invention, may be operatively
associated
With a heterologous nucleotide s~~"pn~.e lrlr_'111d1~1g,hPternl~,g~u~
rvguiatvi~% ri,gi~i2S.iv
i . ~ . ,
increase, decrease, or otherwise modulate, rite expression of a coding region
of
:ntarnet ~zrit~a:~ 1 f ;...,tr....:,...... ,7: ' t
azmvav.m vtmzuz a iiVJl Vl~'GLL11J111. A l.Vlllng ieg1~11W1 Irl~l'.rest may
include, but is not
.
limited to, a gene that encodes a pharmaceutically active protein, for example
growth
factors, growth regulators, antibodies, antigens, their derivatives useful for
immunization or vaccination and the like. Such proteins include, but are not
limited
to, interleukins, insulin, G-CSF, GM-C'SF, hPG-CSF, M-CSF Or ccim__hi_natinnc
thereof, interferons, for example, interferon-a, interferon-13, interferon-T,,
blood
clotting factors, for example, Factor VIII, Factor IX, or tPA or combinations
thereof.
A coding region of interest may also encode an industrial enzyme, protein
supplement, ,
nutraceutical, or a value-added product for feed, food, or both feed and food
use.
Examples of such proteins include, but are not limited to proteases, oxidases,
phytases
chitinases, invertases, lipases, cellulases, xylanases, enzymes involved in
oil
metabolic and biosynthetic pathways etc. A coding region of interest may also
encode
a protein imparting or enhancing herbicide resistance or insect resistance of
a plant
transformed with a construct comprising a constitutive regulatory element as
described herein.
[00127] A list of the nucleotide sequences provided in the present invention
is
provided in Table lb.
57
CA 02507563 2005-05-13
Table lb: Nucleotide Sequence
Summary
.
, ,
. , , . , ,
SEQ ID NO: Name of sequence , ,
1 T 1275 (tCUP) '
' 2 Nde-Sma ,
3 ON '
4 f ~Nm
RENT 1
6 RENT 2
7 RENT 3
,. , ,
~ ~ V T J
9 RENT 7
pr-1 S (primer)
11 pr-3A (primer)
12 pr-2S (primer) .
13 pr-4S (primer)
14 pr-5 A (primer)
pr-6S (primer)
16 pr-7S (primer)
17 pr-8A (primer)
~8
CA 02507563 2005-05-13
. ~ , ~ ~ . c
18 GCC-62-GUS fragment
,
19 HindIII primer ' . ,
20 BglII primer ' ' ~ ' ~ ' ' ' ,
,
~~
21 ' RENT consensus sequence - ~ , '
' , ,
~~ , ,
22 tCUP consensus sequence
, ,,
. ~ ,. , ~ . ,
,~
,
.. , ,
[00128] The present invention will be further illustrated in the following
examples.
Examples ' , '
' ,
Characterization of a Constitutive regulatory element - GUS Fusion
[00129] Transfer of binary constructs to Agrobacterium and leaf disc v ~ .
transformation of N. tabacum SR1 were performed as described by Fobert et al.
(1991, Plant Mol. Biol. 17, 837-851). Plant tissue was maintained on 100
g.glml ,
kanamycin sulfate (Sigma) throughout in vitro culture.
[00130] From the transgenic plants produced, one of these, T1275, was chosen
for detailed study because of its high level and constitutive expression of
GUS.
[00131] Fluorogenic and histological GUS assays were performed according to
Jefferson (Plant Mol. Biol. Rep., 1987, 5, 387-405), as modified by Fobert et
al.
(Plant Mol. Biol., 1991, 17, 837-851). For initial screening, leaves were
harvested
from in vitro grown plantlets. Later nine different tissues: leaf (L), stem
(S), root (R),
anther (A), petal (P), ovary (O), sepal (Se), seeds 10 days post anthesis (S1)
and seeds
20 days post-anthesis (S2), were collected from plants grown in the greenhouse
and
analyzed. For detailed, quantitative analysis of GUS activity, leaf, stem and
root
tissues were collected from kanamycin resistant F 1 progeny grown in vitro.
Floral
59
CA 02507563 2005-05-13
v
tissues were harvested at developmental stages 8-10 (Koltunow et al., 1990,
Plant
Cell 2, 120,1-1224) from the original transgenic plants: Flowers were also
tagged and ' ,
,~
developing seeds were collected from capsules at.10 and 20 dpa. In all cases,
tissue
a ~~ ,
. ,
was weighed, immediately frozen in liquid nitrogen, and stored' at -
80°C. '
~, ,
~,
[00132] ~ Tissues analyzed by histological assay were at the same
developmental ~ ~ , '
stages as those listed above. Different hand-cut. sections were analyzed for
each ' ',
organ. For each plant,, histological assays, were performed on at least two
different , ' ': , ,,
occasions to ensure reproducibility. Except for florar organs, all tissues
were assayed '
, ,
in phoslihate buffer according to Jefferson (1987, Plant ~Llol. Biol. Rep. 5,
387405),
with 1 mM X-Gluc (Sigma) as substrate. Flowexs were assayed in the same buffer
containipg 20% (v!v) methanol (Kosugi et al., 1990, Plarit Sci. 70, 133-140).
[00133], GUS activity in plant T1275 was found in all tissues. Figure 1 shbws
,
the constitutive expression of GUS by histochemical staining with X-Gluc of
T1275,
including leaf (a), stem (b), root (cl, flower (d). ovary (e); e_m_b_rync if
a"rd g)9 and s~PP.d
v
(h). ,
[00134] Constitutive GL'S expression was confirmed with the more sensitive
fluorogenic assay of plant tissue from transformed plant T1275. These results
are
shown in Figure 2. GUS expression was evident in all tissue types including
leaf (L),
stem (S), root (R), anther (A); pistil (P), ovary (O), sepal (Se), seeds at 10
dpa (S 1 )
and 20 dpa (S2). Furthermore, the level of GUS expression is comparable to the
level
of expression in transformed plants containing the constitutive promoter CaMV
35S
in a GUS - nos fusion. As reported by Fobert et al. (1991; Plant Molecular
Biology,
17: 837-851) GUS activity in transformed plants containing pBI121 (Clontech),
which
contains a CaMV 35S - GUS - nos chimeric gene, was as high as 18,770 ~ 2450
(pmole MU per minute per mg protein)
CA 02507563 2005-05-13
Genetic Analysis of Transgenic Plant T1275
[00135) The T-DNA contains a kanamycin resistance gene. Seeds i'rom self
pollinated transgenic plants were surface-sterilized in 70% ethanol for 1 min
and in
undiluted Javex bleach (6% sodium hypochloride) for 25 min. Seeds were then
washed several times with sterile distilled water, dried under laminar flow,
and placed
in Petri dishes containing MSO medium supplemented with 100 ~g/ml kanamycin as
described in Miki et al. (1993, Methods in Plant Molecular Biology and
Biotechnology, Eds., B.R. Glick and J.E. Tompson,'CRC Press, Boca. Raton, 67-
88).
At least 90 plantlets _were counted for each transformant. The number of green
{kanamycin-resistant) and bleached (kanamycin-sensitive) plantiets were
counted a-ft_er
4-6 weeks, and analyzed using the Chit test at a significance level of P<0.05.
[00136) The genetic analysis results are shown below in Table 1 c, which
demonstrates that the T-DNA loci segregated as a single locus of insertion. '
TABLE le: Genetic Analysis of Transgenic Plant T1275
No. of I No. of Observed Expected Chi' I
~
Progeny Progeny Ratio Ratio
~r ~s
262 88 3:1 3:1 0
Consistent
with a single
dominant
gene
Southern Blot Analysis
[00137] The T-DNA in the transgenic plant T1275 was analyzed using either a
GUS gene coding region probe or a nptll gene coding region probe.
[00138) Genomic DNA was isolated from freeze-dried leaves using the
protocol of Sanders et al. (1987, Nucleic Acid Res. 15, 1543-1558). Ten
micrograms
of T1275 DNA was digested for several hours with EcoRI using the appropriate
61
CA 02507563 2005-05-13 ~ ~ , '
. '
~ ~ ~ ~ ,
manufacturer-supplied buffer supplemented with 2.5 mM spermidine. After
electrophoresis through a 0.8% TAE agarose gel, Southern blot analysis was
,, .
conducted using standard protocols. As the T-DNA from the construct containing
the
.; ,
constitutive regulatory element - GUS = nos construct contains only a single
Eco RI
recognition site the hybridizing fragments are composed of both T-DNA and
flanking , ~ ' .
tobacco DNA sequences. The length of the fragment will vary depending on the ~
'
~,
location of the nearest Eco RI site. Using the GUS gene as a probe (Figure 3 -
lane 1),
the fragment to the nearest Eco RI site in the plant DNA will be detected.
With ~ ' '
T 127x, vnc sui:il iagiiient was iUCdieCi. Using ihe~nptli coding region as a
probe,
.: , . , . ., . , ,
(Figure 3 - lane 2), which hybridizes to~ sequences on the opposite side of
the Ecb RI
site, again only one hybridization band, was evident. As'can also be seen in
Figure 3,
no major rearrangements occurred within the T-DNA.
Cloning and Analysis of the Constitutive Regulatory element - GUS Fusion
rnni~m n_._____,_ raT~ ~ , ~ , -
~V~fYJ7 j ~ct~tmtlm ijl~ti vVd~ I~Utai.ed IrOm leaves aCCOrdlng t0 Hattoy et
a~. ' i
(1987, Anal. Biochem. 165, 70-74). Ten ~,g of T1275 total DNA was digested
with
EcoRI and XbaI according to the manufacturer's instructions. The digested DNA
was
size-fractionated on a 0.7% agarose gel. The DNA fragments of about 4 to 6 kb
were
isolated from the gel using the Elu-Quick kit (Schleicher and Schuell) and
ligated to
lambdaGEM-2 arms previously digested with EcoRI and XbaI and phosphatase-
treated. About 40,000 plaques were transferred to a nylon membrane (Hybond,
Amersham) and screened with the 32P-labelled 2kb GUS insert isolated form
pBI121,
essentially as described in Rutledge et al. (1991, Mol. Gen Genet. 229, ~ 1-
40). The
positive clones were isolated. The XbaI-EcoRI fragment (see restriction map
Figure
4) was isolated from the lambda phage and cloned into pTZl9R previously
digested
with XbaI and EcoRI and treated with intestinal calf phosphatase.
[00140] The plant DNA sequence within the clone SEQID NO:1 has not been
previously reported in sequence data bases. It is not observed among diverse
species
as Southern blots did not reveal bands hybridizing with the fragment in
soybean,
62
CA 02507563 2005-05-13
potato, sunflower, Arabidopsis, B. napus, B. oleracea, corn, wheat or black
spruce
(data not shown). In tobacco, Southern blots did not reveal evidence for gross
,
rearrangements at or upstream of the T-DNA insertion site (data not shown).
The T1275 Regulatory Element is Cryptic
[00141 ] The 4.2kb fragment containing about 2.2kb of the T 1275 regulatory '
element fused to the GUS gene and the nos 3' was isolated by digesting.pTZ-
T1275 , .
with HindIII.and EcoRI. The isolated fragment was ligated into the pRD400
vector
;(T~atla vt nl,~~ 1 992 Go;~o~ 211 :3f23=3Q4) pre'vi~rusiy uigeSted with
Hindiii and GcoRi
,,
and treated with calf intestinal phosphatase. Transfer of the binary vector to
Agi~vvaCieriu~ri iuiiiejucien8 and leaf disc transformation of N: tabacum SR1
were
performed as described above. GUS activity was examined in several organs of
many
independent transgenic lines. GUS mRNA was also examined in the same organ by
RNase protection assay (Melton et al, 1984, Nucleic Acids Res. 121: 7035 -
7056)
.
using a probe that mapped the mRNA 5' en_c_1_,'_n tenth ,mtransfn~e.d ~r.~ tr~-
.~~~-.:~
uaAU ~a CLilJ~4i11v
tissues. RNA was isolated from frozen-ground tissues using the TRIZOL Reagent
(Life Technologies) as described by the manufacturer. For each assay 10 - 30
ug of
total RNA was hybridized to an antisense RNA probe as described in Figure 8
(A).
Assays were performed using the RPAII kit (Ambion CA) as described by the
manufacturer. The protected fragments were separated on a 5% Long Ranger
acrylamide (J.J. Baker, N.J.) denaturing gel which was dried and exposed to
Kodak X-
RP film.
[00142] RNase protection assays performed with RNA from leaves, stem, root,
developing seeds and flowers of transgenic tobacco revealed a single protected
fragment in all organs indicating a single transcription start site that was
the same in
each organ, whereas RNA from untransformed tobacco tissues did not reveal a
protected fragment (Figure 8 (B)). The insertion site, including 1200 by
downstream,
was cloned from untransformed tobacco as a PCR fragment and sequenced. A
composite restriction map of the insertion site was assembled as shown in
Figure 8
63
CA 02507563 2005-05-13 ~ '
,
,~ , ;
(A). RNA probes were prepared that spanned the entire region as shown in
Figure 8
(A). RNase protection assays did not reveal transcripts from the sense strand
as ~ '
summarized. in Table 2. These data suggest that the insertion site is
transcriptionally
,a ~ ~ ,
silent in untransformed tobacco and is activated by T-DNA insertion. The
~egion '
upstream of the insertion site is therefore anpther exarraple of a plant
cryptic regulatory , '
. ~ '
element. , '
~, ,
Table 2. Summary of the RNase Protection Assays of the insertion site in ~ , '
1; , ,,
.
untransfot~med tobacco. See Figure 8'(A) for probe positions. ~ , ,
., ,
Probe Rnase Protection Assay result ' '
Looicing,for "sense" RNAs (relative,.to the T1~75 regulatory element)
C8-EcoRI many bands, all in tRNA (negative control)
A10-HindIII no bands 1 ~ ~ ,
2-21-HindIII no bands '
1-4 ~m_~I manor hande alb i,~ tT?AT n ' ~ .
.:~ . tauvr
i I ~.
7-EcoRI faint bands, all in ~tRNA ,
Constitutive Activity of the T1275 Regulatory Element
[00143] For analysis of transient expression of GUS activity mediated by
biolistics (Sandford et al, 1983, Methods Enzymol, 217: 483-509), the Xbal -
EcoRl
fragment was subcloned in pUC 19 and GUS activity was detected by staining
with X-
Gluc as described above. Leaf tissue of greenhouse-grown plants or cell
suspension
cultures were examined for the number of blue spots that stained. As shown in
Table
3, the T1275 - GUS nos gene was active in each of the diverse species examined
and
can direct expression of a coding region of interest in all plant species
tested. Leaf
tissue of canola, tobacco, soybean, alfalfa, pea, Arabidopsis, potato,
Ginseng, peach,
and cell suspensions of oat, corn, wheat, barley and white spruce exhibited
GUS-
positive blue spots after transient bombardment-mediated assays and
histochemical
64
CA 02507563 2005-05-13 .
GUS activity staining. This suggests that the T1275~ regulatory element may be
useful
. for directing gene expression in both dicot and monbcot pants. , ~ , ,
TABLE 3: ~ Transient Expression of GUS Activity in Tissues of Diverse Plant
Species , ,
Tissue Source Species . GUS Activity
Leaf Soybean +++
. , Alfalfa ~ , ++
,
, , Arabidopsis ~ , ~ . +
. ' I Potato ++
f
Ginseng. ~ ~ ++
~
Peach ' + !
Leaf disc Tobacco , ++ '
B. napes +
Pea ~ ~ +
Cell Cultures Oat +
Corn +
W_h_Pat +
Barley , ++
White spruce ++
* Numbers of blue spots: 1 - 10 (+), 10 - 100 (++), 100 - 400 (+++)
[00144] For analysis of GUS expression in different organs, lines derived from
.
progeny ef tfe above lines were examined in detail. Table 4 shows the GUS
specific
activities in one of these plants. It is expressed in leaf, stem, root,
developing seeds
and the floral organs, sepals, petals, anthers, pistils and ovaries at varying
levels,
confirming constitutive expression. Introduction of the same vector into B.
napes also
revealed expression of GUS activity in these organs (data not shown)
indicating that
constitutive expression was not specific to tobacco. Examination of GUS mRNA
in
the tobacco organs showed that the transcription start sites were similar
(Figure 8 (B)),
and the level of mRNA was similar except in flower buds where it was lower
(Table
4).
CA 02507563 2005-05-13
~. ,
TABLE 4: GUS Specific Activity and Relative RNA Levels in the Organs of
Progeny of Transgenic Line T64 ' , , ~ , ,
Organ Relative GUS GUS Specific
RNA Activity
Levels in T64 (picomollMU/min/mg
protein)
Progeny (grey ~ ,
scale
' units) .
TransformedUntransformed
Tobacco Tobacco .
T64
Leaf , 1774 988.32 ,, 3.02
,
' Steiu ' i82v 826.48 ~ 7.58 , ,
.. ~
Root 1636 ~ 4078.45 22.18
T '
14 day post l 790 253.21 y 10.03 '
anthesis Seeds
Flower - buds715 2.59 ~ ND*
Petals ND* 28.24 1.29
~
n - 4 rti,nro TvTTI* ~ r n .
~.o~ i~.33
Pistils ND* 9.76 ' 1.72
Sepals ND* 110.02 I 2,48
Ovary ND* 4.42 2.71
* Not Done
T1275 sequence comparison
[00145] The present invention provides an isolated nucleotide sequence
selected from the group consisting of SEQ ID NO: 1, and a nucleotide sequence
that
hybridizes to SEQ ID NO:l under a condition selected from the group consisting
of:
- hybridizing overnight (16-20hrs) in a solution comprising 7% SDS, O.SM
NaP04 buffer at pH 7.2, and 10 mM EDTA at 65°C and washing for one hour
at 60°C
in a solution comprising 0.1 X SSC and 0.1% SDS;
66
CA 02507563 2005-05-13
~. ' ,
- hybridizing overnight (16-20hrs) in a solution comprising 7% SDS, 0.5M
NaP04 buffer at pH 7.2, and 10 mM EDTA at 65°C and washing for one hour
at 65°C ' ,
in a solution comprising 2 X SSC and 0.1% SDS; and"
.,
- hybridizing overnight (16-20hrs) in a solution comprising 4 X SSC at
65°C ,
and washing for one hour in 0.1 X SSC at 65°C;
~, ~ , . ,
wherein the nucleotide sequence confers constitutive expression of a coding
region of '
interest linked in operative association therewith. , ' '
. . , ,
,, ,
[OOi46~' The Tm of T1275 is compared to the clos2st homoi_og~ae idPr!tifPd In
a '
sequence similarity search. an At-abid~nsi.c n_h_~~rn~helas_,'_,_.mthase be a
(~=a~ha~W
i v aav
r ~ ,
Acc~ssian No.AF085230) that exhibits 52% similarity with T1275. The following
analysis indicates that the T 1275 sequence and' nucleic acid sequences that
hybridize
to T1275 under stringent conditions defined herein are unique.
[00147 T he T m°C, under the hybriaization conditions stated above ~for
T1275 ;
and AF085230, are provided in table 4A below, where,
- % similarity was calculated using NCBI Blast 2 program (available through ,
the NIH at: ncbi.nlm.nih.gov/cgi-bin/BLAST/; parameters for alignments were
set at:
match 5; mismatch -4; gap open 5; gap extension 2; x dropoff 50; expect 10;
worksize 11 and filter ON).
- Conditions for A, B and C listed in Table 4A are:
A= 7% SDS, O.SM NaP04, lOmM EDTA, at 65°C (hybridization)
B= 2xSSC, 0.1%SDS, at 65°C (washing)
C= O.IxSSC, o.l%SDS, at 60° or 65°C (washing)
67
CA 02507563 2005-05-13 ,
- Tm~(perfect match) is calculated using the formula described in Baldino et
al
(Baldino, Chesselet and Lewis 1989. High-resolution in situ hybridization.
histochemistry. Methods in Enzymology 168: 761-777) using the
following~formula:
Tm = 81.5 + 16.6(log[Na+]) + 0.41(%G+C) - 675/(PL) - 0.65(%formamide).
-PL is the probe length in bases;
-Tin (heterologous match) was calculated by the method as described in
Baldino et ~l (Baldino, Chesselet and Lewis 1989'. High-resolution imsitu
' ,
' hybridization histochemistry. Methods in Enzyrnohg~ 16R: 761-777) and~Borner
et al
(Bonner, Brenner, Neufeld and Britten 1973. Reduction in the rate of DNA
reassociation by sequence divergence. J. Mol. Biol. 81: 123-13S), using the
following
formula:
Tm (heterologous match) = Tm(perfect match)-1.0(%mismatches, including gaps).,
TabIQ 4A
nucleotide sequence of % similarity with Tm °C Tm °C
SEQ ID NO:1 AF085230 (perfect match) (heterologous match)
A B C A B C
1-2224 52 79 73 53 31 . 25 5
[00148] These results of the above calculations show that there is about 1
degree C of decrease in Tm for each % mismatch between two DNA sequences.
Assuming a perfect match (100% similarity, which is not the case) between the
sequence disclosed in AF085230 and that of SEQ ID NO:1, the results shown in
Table
4A demonstrate that a Tm of less than 79°, 73°, and 53°C
is required to detect ,
hybridization between nucleotides 1-2224 of SEQ ID NO:1 and the sequence of
AF085230 under the hybridization and washing conditions stated above.
68
CA 02507563 2005-05-13 '
~ . ~. , s
Furthermore, taking into account the % similarity between nucleotides 1-2224
of SEQ.
ID NO:1 and the sequence of AF085230, the results in Table 4A demonstrate that
a.
hybridization temperature of greater than 31 °C,(Tm heterologous match)
will not
. ,
result in hybridization between nucleotides 1-2224 of SEQ ID NO:1 and
AF085230. ' ' '
. , , ~, , .
~, ' ,
[00149] ~ As the temperatures stated for hybridization above are from
60° to ~ , '
' ~ ,
65°C, and are well above the calculated Tm's indicated in Table 4A,
abbve,~the , ;
hybridization conditions stated for T1275 do not detect the nucleotide
sequence ' ,
comprising AF085230. Therefore, the T1275 sequence and riucleiclacid,sequences
'
.. .
that hybridize to T1275 under stringent conditions fdefin~d'herein are uni
fue. ~ , ' '
I~lenti~catio'n of Regulatory Elements within the Full ~,ength T1Z75
Regulatory
Element
[00150] An array of deletions of the full length regulatory region of T1275
were '
prepared, as identified in Figures 5 (A) and (B), far further analysis of the
cryptic .
regulatory element. ' ~ , ,
Plasmid Construction
[00151 ] Deletion and replacement constructs were created in the vector pBI221
(Clontech), which contains the GUS (uidA) coding region driven by the CaMV 35S
promoter and the NOS terminator. Independent constructs representing 5'
deletions of
the tCUP were generated at convenient restriction sites within the tCUP
sequence.
The CaMV 35S promoter of pBI221 was replaced with the deletion fragments of
tCUP to generate -1304-GUS, -684-GUS, -394-GUS, -197-GUS and -62-GUS. The
numbers represent the nucleotide numbers relative to the transcription
initiation site.
[00152] Fragments to test the enhancer elements between the fragments -394 to
-62 (1660-1992 of SEQ ID N0:1 or 22) and -197 to -62 (1875-1992 of SEQ ID NO:l
or 22) relative to the transcription start site of the tCUP were amplified by
PCR with
69
CA 02507563 2005-05-13
. Taq DNA polymerase. The fragment from -394 to -62 was amplified with pr-1
and pr- '
. 3 primers: ~ , , ~ ,
pr-1 S: TTGCCTGCAGGGGATCTTCTGCAAGCATC (SEQ ID NO:10); and '
. pr-31A: TCAAATGCATGGATCAAAAGGGGAAAC (SEQ ID NO:11 ),
and the fragment from -197 to -62 was amplified with pr-2
. , , pr-2 S: GGAGCTGCAGGCTATTTAAATACTAGCC (SEQ ID N0:12) and
~~
Ipr-3 primers. All primers had additional nucleotides at the 5' ends to gi~,re
ahe Pstl '
restriction sites for subcloning PCR products. The PAR products were ligated
into the
Pstl sites located upstream of the -394-GUS and -197-GUS to generate -394(2X)-
GUS and -197(2X)-GUS constructs. ~ ,
[00153] A ~6 minimal 35S promoter (-46-35S) was generated by PCR using
' 1 the pr-4 and pr-5 primers: .
pr-4 S: CACTCTGCAGGCAAGACCCTTCCTCTATA (SEQ ID N0:13);
pr-5 A: ATATAAGCTTTGGGGTTTCTACAGGACG (SEQ ID N0:14))
and pBI221 DNA as a template. The PCR product was digested with Pstl and
BamHI,
and the resulting fragment was used to replace the PstlI and BamHI fragment in
pBI221. The fragment from -197 to -62 of tCUP (nucleotides 1875-1992 of SEQ ID
NO:1 or 22) was subcloned into the the PstI sites located upstream of the -~6-
35 S-
GUS to generate -197-35S-GUS, -1978-35S-GUS ,and -197(2X)-35S-GUS
constructs.
?0
CA 02507563 2005-05-13
i,
s
[00154] ~ The -12-GUS construct was generated by PCR using the pr-6 and pr-5 .
primers: ,
,~ ,
pr-5 A: ATATAAGCTTTGGGGTTTCTACAGGACG,(SEQ iDN0:14); ,
pr-6 S: GAGAAGATCTCCAAACACCCCT .AACTCTATC (SEQ ID'NO:15). ,
[00155] The PCR product was digested with XbaI and KpnI, and the resulting
fragment was used to replace the XbaI.and KpnI fragment in tCUP-GUS. To
generate , ' ' ;
the -62-tsr=GUS construct, the DNA sequence bet~~en~=62' and -12 of tCUP was
amplified with the pr-7 and pr-8 primers: ~ ~ ~ ' ' ' , ' ~ ,
pr-7 S: TTGATCATT TTGATCAACGCCCAG (SEQ ID N0:16);
pr-8 A: AGGGGGTGCATATGAATTAAAAAAGGAAAAG (SEQ ID ,
N0:17). ,
[00156] The PCR product was digested with XbaI and NdeI, and the resulting
fragment was used to replace the XbaI and IVdeT fi-a'gme_n_t in r_C'.LTp-Cv'TI
rS~, 'f he T A 3~_ ~
GUS construct was generated using pr-9 and pr-5 primers. To generate GCC-62-
GUS
construct, a 51-by fragment: '
GCC-62-GUS fragment:
GCATAAGAGCCGCCACTAAAATAAGACCGATCAAATAAGAGCCGCCATG
CA (SEQ ID N0:18)
containing two GCC boxes (GCCGCC; Ohme-Takagi and Shinshi, 1995, Ethylene-
inducible DNA binding proteins that interact with an ethylene- responsive
element.
Plant Cell 7: 173-182) was ligated into the PstI site located upstream of the -
62-GUS
construct.
71
CA 02507563 2005-05-13
. i'
, . ,
~, , , s
Plant Transformation and Selection
[00157] ~ ' , Ardbidopsis thaliana (ecotype Columbia) was grown in a growth
chamber (16 hr of light and 8 hr of darkness at ~23°C) after, a 2-4 day
~ernalization ,
period. For growth under sterile conditions,. seeds were ,surface 'sterilized
(15 min', , '
~~ '
incubation in 5% [v/v] sodium hypochlorite, and a three-time rinse in sterile
distilled , ~ '
water) and sown on half strength Murashige and,Skoog salts (Sigma),
supplemented
with 1 % sucrose, pH 5.7, and 0:8% (w/v)' agar in Petri dishes. ~ ~ , ~ ~ ; ,
,,
. ,
[00158] All the consticuts and GUS fusion vi~ere subcloned into the pRD400 ,
,
(Latla RS, Harnrneriindi 3K, Panchuk is, Pelcher LE, l~eller W: Modified
binary plant
transformaticn vectors with the wild-type gene encoding ~1PTII. Gene 211: 383-
384,
1992) or pCANIBIA2300 (Cambia, Canberra, Australia) binary vectors for plant.
transformation. Plant transformation plasmids were electroporated into
Agrobac~terium
tumefaciens GV3101 (Van Lar~beke, N, Engler, G, Holsters, M, Van den Elscker,
S,
Zainen, I, Schilperoort, RA, and Schell, J:. Large plasmid in Agrobacterium
tumefaciens essential for crown gall-inducing ability. Nature 252,169-170,'
1974) as''
described by Sha:v (Sha~t~ CH: Introductory of cloning plasmids into
Agrobacterium
tumefaciens. Meth Mol Biol 49, 33-37, 1995). The Agrobacterium-mediated ,
transformation of Arabidopsis thaliana was performed as described (Clough SJ,
Bent
AF: Floral dip: a simplified method for Agrobacterium-mediated transformation
of
Arabidopsis thaliana. Plant J. 16: 735-743, 1998,), with the following
modifications.
Plants with immature floral buds and few siliques were dipped into a solution
containing Agrobacterium tumefaciens, 2.3 g/L MS salts (Sigma), 5% (wlv)
sucrose
and 0.03% Silwet L-77 (Lehle Seeds, Round Rock, TX) for 0.5 min. Tl seeds were
collected, dried at 25°C, and sown on sterile media containing 40~gJmL
kanamycin to
select the transformants. Surviving T1 plantlets were transferred to soil. 15
to 30
independent transgenic lines for each construct were selected and used for the
analysis.
of GUS activity.
72
CA 02507563 2005-05-13
Re~ulatory element activity in tomato: protoplast isolation and
electroporation
[00159] Young and fully expanded leaves were excised from ,about 4 weeks old .
tomato plants and surface sterilized in 5% commercial bleach (Javex) (1%
NaOCI).
The abaxial surface of leaves were gently rubbed with earborandum powder and
rinsed three times with sterile water. After removing midribs, the~remaining
leaf
blades were cut by sharp razor into small pieces and 'floated on enzyme
mixture
containing 0.3% Cellulase Onozuka R-10 (Yakult Honsha), 0.15% macerozyme R-10
(Yakult Horisha) and 0.4 M sucrose.
[00160] After overnight incubation in dark at 30°C, protoplasts were
collected
by filtration through a 100~m nylon mesh filter followed by centrifugation at
500 rpm
for 5 min. The floated protoplasts were gently collected by a wide bore
pipette and
washed twice with electroporation buffer (150 mM ~KCI and 0.4 M mannitol) for
5
min at 400 rpm and finally suspended at approximately 1x106Jm1 in O.S M
mannitol
containing 150 ~M MgCl2.
[OU 161 ] 'fhe viability of protoplasts was confirmed by fluorescin diacetate
and
alanine blue staining and protoplasts were kept on ice for 30 minutes prior to
electroporation. A 25-30 ~g plasmid DNA (see Figure 5 (H) for added
constructs)
was added to 5008.1 protoplast syspension, mixed gntly and electroporated at
100~.F
and 200 Volts using Gene Pulser II (BioRad). To normalize for transfection
efficiency, the CaMV 35S promoter-luciferase plasmid was cotransfected in each
experiment. The electroporated protoplasts were kept on ice for 15-30 min,
centrifuged for 5 min at 500 rpm and mixed with 0.5 ml Murashige and Skoog
medium (containing 3% sucrose, 9% mannitol, 0.1 mM MgS04, 2 mg/L
naphthylacetic acid and 0.5 mg.~I, benzyladenine). The cultures were kept in
dark at
25°C for 24 hr, and cells were collected in microcentrifuge tubes. To
each 500p.1 of
protoplast suspension 200 ~1 of buffer solution (100 mM sodium phosphate, pH
7.8, 1
mM EDTA, 0.5% Triton X-100, 70 mM 2-mercaptoethanol and 10% glycerol) was
added and protoplasts were lysed for lucerifase and GUS assay.
73
CA 02507563 2005-05-13 '
,~ ,
Deletion analysis of tCUP , ,
[00162] ~ ' , In order to delineate functi~b~nal regions, of the tCUP'
regulatory, a series
of 5' deletion constructs were made (Figure's (J)), anc~~ activities
were'examined in ,
,:
leaves of transgenic Arabidopsis plants. As shown in Figure 5 (K) and ,Table 5
below, , '
' . ~ ' ,
all sequences from about -2054 to ~ -684 (nucleotides about 290 to 1370 of SEQ
ID ~ , '
NO:1 or 22) relative to the transcription initiation site of the tCUP promoter
could be ' ' .
deleted with no significant effect on promoter activity. Deletion of sequences
to'-394 ~ ~;
. ,
and -197 (nucleotides 1660-1875 of SEQ ID NO:1 at 22) dbcreased expression
about
40% and 60%, respectively. The -62 deletion construct reduced GUS~activity td
a~ ~ .
,
level slightly over background. These results indicated that the -62 fragment
contained
the minimal promoter and positive~cis'~regulatary elements were potentially
located in
the regions from:
-684 to -394 (nucleotides 1370-1660 of SEQ ID NO: l or 22);
-394 to -197 (nucleotides1660-1875 of SEQ ID NO:1 or 22); and ~ ;
-197 to -62 {nucleotides 1875-1992 of SEQ ID.NO: i or 22).
Identification of enhancer elements
[00163] To locate enhancer activities within the fragments -394 to -62
(nucleotides 1660-1992 of SEQ ID NO:1 or 22),~and -197 to --62 (nucleotides
1875-
1992 of SEQ ID NO:1 or 22), these fragments were duplicated in the promoter
constructs, -394(2X)-GUS and -197(2X)-GUS (Figure (J)), and GUS activity was
analyzed in transgenic Arabidopsis plants. As shown in Figure S (K), insertion
of two
copies, of -197 to -62 and -394 to -62 fragments (nucleotides 1660-1992 and
1875-
1992, respectively, of SEQ ID NO:1 or 22) increased promoter activity about
1.5 to 2-
fold compared with the constructs with only one copy of these fragments.
[00164] To evaluate whether the enhancers within fragment -197 to -62
(nucleotides 1875-1992 of SEQ ID NO: l or 22) could function with other core
74
CA 02507563 2005-05-13
promoters, the fragment was also fused to the -46 minimal promoter of CaMV 35S
(Figure 5 (L)). As shown in Figure 5 (M), insertion of one copy the, fragment
in bpth
the forward. and reverse orientation increased GUSI activity by about 15-fold
in leaves
' of transgenic Arabisopsis. Insertion of two copies further enhanced GUS
activity by .
40-fold. This suggests that the fragment -197 to -62 (nucleotides ~ 875-1992
of.SEQ
ID NO:1 or 22) may function as a transcriptional enhancer element.
Analysis of core promoter region
[00165] ~ To analyze the tCUP core promoter, a series of deletions or
~r~odificatiorm surrounding tire transcriptionai start site were made (Figure
5 (H)). ' I
Promoter activities were examined using a transient assay in tomato
protoplasts
(Figure 5 (I)):
- deletion of the core promoter to -12 (position 2042 of SEQ IDNO:1 or 24)
decreased GUS activity by 40%;
- deletion of the sequence surroundings the transcription start site reduced
it tb
2°,% of the -62-GT~JS construct activity, suggesting that the
transcription start site
sequence was essential for tCUP promoter activity;
- substitution of the sequence -30 to -24 with a TATA-box (TATATAA) in
the -62-GUS construct increased the promoter activity about 3-fold;
- addition of GCC-box sequences (Hart CM, Nagy F and Meins Jr F: A 61 by
enhancer element of the tobacco beta-1,3-glucanase B gene interacts with one
or more
regulated nuclear proteins. Plant Mol Biol 21, 121=131,1993; Ohme-Takagi M,
Shinshi H: Ethylene-inducible DNA binding proteins that interact with an
ethylene-
responsive element. Plant Cell 7: 173-182,1995) further increased the core
promoter
activity to about 4-fold.
[00166] 5' deletions of the regulatory element (see Figures S(A) and (B) and
analysis by transient expression using biolistics showed that the regulatory
element
7~
CA 02507563 2005-05-13
was active within a fragment 62bp from the transcriptional start site
(position 1992 of .
SEQ ID NO:1 or 22) indicating that the core promoter'has a basal level of
expression
., ,
(see Table 5: Figures 5 (D) and (I)). ~ . " .
:~ ,
', ~ ~ ,
Table 5: Transient GUS activity detected. in soybean, leaves by staining with
X-
, ."
gluc after particle bombardment. Vectors illustrated in Figures. 5 (A) and
(B). , ~ , '
Genes (nucleotides) , ,GUS staining '
SEQ ID NO:1 or 22 ' '
1. T1275-GUS-nos (1-2224). . ~ , ,~- ' . ,
2. ' -1639-GUS-nos (705-2224) ~ , ~ + , , . , , , ' , .
3. -1304-GUS-nos (1040-222x) ' ~ + ' , ' ' ,.
4. -684-GUS-nos (1370-2224) ' , +
5,. -394-GUS=nos (1660-2224) ' ' + ~ , ,
6. -197-GUS-nos (1875-2224) , +
7. -62-GUS-nos (1992-2224) ~ ~
8. -62(-tsr)-GUS-nos + ' '
9. -12-GUS-nos (2042-2224) + , '
10. +30-GUS-nos - ' -
~~ ,
[00167] Deletion of a fragment containing the ~transcriptional start site (see
-
62(-tsr)/GUS/nos in Figure 6(B), Table 5) did not eliminate expression,
however
deletions to +30 (+30-GUS nos) reduced expression dramatically. Similar
results
were obsereved in transgenic tomato (see below; Figures 5 (H) and (I))
indicating that
the region defined from about -12 to about +30 contained the core promoter.
DNA
sequence searches did not reveal conventional core promoter motifs within this
region
as are typically found in plant genes, such as the TATA box.
[00168] Deletion of a fragment containing the transcriptional start site (see -
62(-tsr)-GUS-nos in Figures 5 (B), (H) and (I); Table 5, Examples) reduced
expression dramatically in transgenic tomato, however deletions to +30 did
eliminate
expression indicating that the region defined from about -12 to about +30 by
contained the core promoter. Deletion of sequences surrounding the
trasncriptional
start site, reduced activity to about 2% of the activity associated with the -
62-GUS
76
CA 02507563 2005-05-13
' construct, indicating that the trasncriptional start site sequence' is
required for tCUP '
' regulatory element activity. ' , ~ , ,
[00169] ~ A number of the 5' regulatory element deletion clones (Figure 5(A))
were transferred into tobacco by Agrobacterium-mediated transformation using
the
' vector pRD400. Analysis of GUS specific activity in leaves of transgenic
plants (see
Table ~6) confirmed the transient expression data dovim to the -19.7 fragment
' (nucleotide 1857 of SEQ ID NO:1). ' ,
Table 6: GUS specific activities in leaves of greenhouse-grown transgenic
iuiOacCO, Sici, transformed with the T iz75-GUS-rios gene fusion ar_d 5'
deletion
clanes (see Figure 5 C). MeantSE(n)
Genes nucleotides GUS specific activities
SEQ ID NO:1 or 22 pmoles MU/min/mg protein
1.T1275-GUS-nos(1-2224) 283.1171 (27)
2.-1639-GUS-nos(705-2224) 587188 (26) ,
,
3.-1304-GUS-nos(1040-2224) 632217 (10)
4.-b84-GUS-nos (i3r0-2224) not determined
S.-394-GUS-nos (1660-2224) 1627340 (13)
6.-197-GUS-nos (1875-2224) 47574 (27)
[00170] Histochemical analysis of organs sampled from the transgenic plants
indicated GUS expression in leaf, seeds and flowers. .
[00171 ] Deletions in the upstream region indicate that negative regulatory
elements and enhancer sequences exist within the full length regulatory
region.
Deletion of the 5' region to BstYI (-394 relative to the transcriptional start
site)
resulted in a 3 to 8 fold increase in expression of the gene associated
therewith (Table
6), indicating the occurrence of at least one negative regulatory element
within the
XbaI-BstYI portion of the full length regulatory element. Other negative
regulatory
elements also exist within the XbaI- BstYI fragment as removal of an XbaI-PstI
fragment also resulted in increased activity (-1304-GUS-nos; Table 6).
77
CA 02507563 2005-05-13
[40172] ~ To determine if enhancer elements exist, fragments -394 to -62
(nucleotides 1660 to 1992 of SEQ ID NO:I) and -197~to -62 (nucleotides 1875 to
,
,. '
1992 of SEQ ID NO:1) were fused to the -46 35S core promoter. Both fragments
' ' ' raised the expression of the core promoter about 150 fold (Figure 5 (D),
constructs
DRAT-35S and BSTl-35S). Doubling of the -394 to -62 region (nucleotides 1660
to
1992 of SEQ ID NO:1 ) resulted in a 1.8 fold increase in GUS activity when
fused to
T1275 core promoter (BST1-GUS (-394-GUS) v. BST2-GUS; Figure 5~(D)), a similar
,
effect is observed when the -394 to -62~region is double and fused to the 35S
core
' ~ , promoter (BSTl-35S v. BST2-35S). Doubling of the -197 to -62 fragment
(:~acleotides 1875 to i 992 of SEQ iD i3O: i ) aiso produced increased GUS
activity
when fused to the T1275 core promoter (DRA2-GUS). ,
[00173] The -197 to -62 fragment (nucleotidcs 1875 to 1992 of SEQ ID NO:I; '
DRA1-35S), the -197 to -62 fragment in reverse orientation, o_r inverted (DRA
1R-
35S), and a repeat of the -197 to -62 fragment (DRA2-35S) were also fused with
the .
35S minimal promoter (Figure 5 (E) and used to transform Arabidopsis:
[00174] A,°abidcpsis plants wi'~h immatwe floral buds and few silques
were
transformed with the above constructs by dipping the plant into a solution
containing
Agrobacterium tumefaciens, 2.3 g/L MS, 5% (w/v) sucrose and 0.03% Silwet L-77
(Lehle Seeds, Round Rock, TX) for 1-2 min, and allowing the plants to grow and
set
seed. Seeds from mature plants were collected, dried at 25°C, and sown
on sterile
media containing 40p,g/mL kanamycin to select transformants. Surviving
plantlets
were transferred to soil, grown and seed collected.
[00175] Constructs comprising the -197 to -62 fragment (nucleotides 1875 to
1992 of SEQ ID NO:1) in regular or inverted orientation exhibited increased
transcriptional enhancer activity, over that of the minimal promoter (Figure ~
(F). A
further increase in activity was observed when plants were transformed with
constructs comprising repeated regions of this regulatory element (Figure 5
(F).
Tissue staining of transformed plants expressing DRAT-35S indicated that this
78
CA 02507563 2005-05-13
construct was expressed constitutively as it was detected in all tested
organs, including
. ,
flower, silque and seedling (Figure 5 (G)). '
RENT (Repetitive Element from N tabacum) family of repetitive'elements ,
[00176] An amplified N. tabacum line SRl custom library (Stratagene), which .
;
contained MboI partially digested genomic DNA in' the 8-DashII vector, was
screened
'
by hybridization with 32P-labelled probe fragin~nt S (probe 5 is a BstYI- SmaI
,
~ , ,,
fragment of T1275, nucleotides 1660-2224 of SEQ Ip NO:,I, see frgure-5 (C)) at
65°C
~. ,
over ''night (16-20 hours in Churches buffer: 7% fDS; O.,SM NaP04; IOmM EDTA)
,
and washing at 50°C in 0.1 X SSC, O.j% SDS for 60 minutes, or two
washes of 20-30
minutes.each. Approximately 70 clones were identified in this manner. The ,
restriction fragment of each 8 insert which hybridized with probe fragment 5
on a
Southern blot, hybridized at 65°C (overnight; 16-20 hours) and washed
at 60°C,10.1 X
SSC, 0.1% SDS (for 60 minutes, or two washes of 20-30 minutes each; stringent
hybridization conditions), was gel purified with EluQuick (Schleicher and
Schuell) ,
and subcloned _into pGEM4Z (Promegal. Rot_h_'strands of Pach s,~bclone ~=.~ere
sequenced with universal or custom designed primers, as appropriate. From this
screen, 5 clones were obtained for further analysis. Approximately 2 to 3 kb
of each '
genomic clone was subcloned and overlapping sequences obtained. These clones
are
called RENT 1, 2, 3, 5 and 7.
[00177] Two primers, approximately 30 basepairs in length were synthesized
(Synthaid Biotechnologies Inc.), one in the forward direction at position 1707
of the
T1275 nucleotide sequence and the other in the reverse direction at position
2092.
Each incorporated a convenient restriction site, the first a HindIII site:
HindIII primer: TTA AGA TTT AAT Taa get tAT AAT TAC AAA (SEQ ID N0:19)
and the second a BgIII site:
BgIII primer: ATT Cag atc tGG CGG TTGGTG AGA AA (SEQ ID N0:20).
79
CA 02507563 2005-05-13
' [00178] ~ The primers were then used for PCR~ amplification of each of the
five
RENT fragments with attached restriction sites using ~Taq, DNA polymerase
(frorrx
MBI Fermentas Inc). The protocol accompanying the modifying enzyme was ~ ,
' ' ' followed, with a reduction to. 0.2u1 in the amount of Taq DNA polymerase
used, in a ,
total reaction mix of 50w1. The fragment from the original T127~ sequence was
also
amplified.
[00179] All PCR products were,electrophoresed on a 1% TAE agaxose gel and
visualized fy staining with ethidium bromide. The ;400 basepair band
representing the
PCR product was excised and purified. Each DNA,sample was then digested with '
Hind IIL~Bgl II and concentrated in an overnight precipitation with one half
volume of
7.5M ammonium acetate and 2 volumes of 95% ethanol.
[00180] A plasmid containing the vector, pTZl9R, containing the tCUP delta
,,
regulatory element, with a Kozak sequence was also digested with Hind III/Bgl
II, '
electrophoresed on a 1 % agarose gel and gel purified. Briefly, tCUP delta
(see below,
description relating to Table 10 and Figure 10) was 'created by replacing the
NdeI site
(Figure 10(A)) within the leader sequence to a Bglii site thereby eliminating
the
upstream ATG at position 2087 of SEQ ID NO:1. A Kozak consensus sequence was
also constructed at the initiator MET codon and a NcoI site was added to
facilitate
construction with other coding regions (see Figure 10 (B)). Nucleotides 1-86
of SEQ
ID N0:3 (i.e. tCUP delta with Kozack sequence) are derived from T1275
(nucleotides
2086-2170 of SEQ ID NO:1), and a Kozack sequence from nucleotides 87 to 97 of
SEQ ID N0:3. Nucleotides 98 to 126 of SEQ ID N0:3 comprise the vector sequence
between the enhancer fragment and the GUS ATG. The GUS ATG is located at
nucleotides 127-129 of SEQ ID N0:3.
[00181] Each of the five RENT PCR fragments, as well as the T1275 control
PCR fragment was ligated into the digested plasmid, in a 4 to 1, insert to
vector ratio.
These were transformed into ToplO competent cells (Invitrogen Corp.) via
electroporation using an Invitrogen electroporator and their supplied
protocol. The
CA 02507563 2005-05-13 '
w , i
transformed cells were plated on ampicillin containing LB plates and
allowedlto grow. '
overnight. The colonies were then grown overnight irr liquid LB plus
ampicillin to be ' ,
used for plasmid isolation using the Wizard Plasmid Miniprep Kit,(Promega
Corp.) or
.
the Qiaprep Spin Miniprep Kit (Qiagen Inc.). Isolated plasmid5 were restricted
with '
Hind III, Bgl II and Hind III/Xba I to verify restriction ,patterns. Once
these were . . , '
ascertained to be correct, the insert containing plasrxiids were sequenced.
Therefore,
the regulatory elements used fox the analysis in Figure 14 (A), including~tCUP-
RENT, ~ '
~~~ , ~ n _
consist of the amplified PCR fragment fused to tCUP delta comprising a, Kozak
~ ''
'. ~ ,
sequence. The 35S-46 construct used for the analysis presented in Figure 14
(A) was
prepared by generating a -46 minimal 3~SS proineter (-46-3JS) was g2rlGralGd
by ~PCR
using the primer pair: ~ . ',
46-35S-1 primer: CACTCTGCAGGCAAGACCCTT~CTCTATA (SEQ ID N~:13),
' ,
and '
ATATAAGCTTTGGGGTTTCTt~CAGGACG (SEQ ID NO:14),
and pRI221 (lv'1C itech) Dl'VA as a te~iyiate. T iie PCR product was digested
with Pstl
and BamHI, and the resulting fragment was used to replace the PstII and BamHI
fragment in pBI221.
[00182] Approximately 2 to 3 kb region of each genomic clone, which on
Southern blots hybridized with probe 5 (a BstYI-~'maI fragment) was subcloned
and
overlapping sequence reads were obtained on both DNA strands of each subclone.
Sequence analysis indicated the presence of sequence similarity, but not
identity,
along the 3' ends of these subclones, with divergence at the 5' ends. The 5'
ends of the
clones all diverged at the same position. These data suggest that each
independent
clone represented a different member of the RENT repetitive element family
interrupting different regions of the genome. Moreover, all five subclones
studied
were similar to the tCUP sequence in the region which delimits maximal
regulatory
element activity and is situated towards one end of RENT. The five subclones
exhibited 77 to 92 % (Figures 13 (A) - (C)) with sequence similarity with the
tCUP
81
CA 02507563 2005-05-13
sequence in the probe 5 region (1724-2224 of SEQ ID NO:1) which confers
regulatory
element activity. The repetitive elements also do nat appear to be pxesent in
close,
tandem locations as probe five hybridized only once v~itf~ each genomic close.
[00183] Therefore, t-CLJ~ is a member of a large family of repetitive elements
in Nicotiana tabacum (RENT) in which the regions essential for regulatory
element
activity have been conserved. All RENT sequences,~including t~UP share a
common
,.
sequence of ca. 525 by from transcriptional start site of t-CUP (1724-2224 of
SEQ ID
' ,
NO:1). RENT sequences 1, 2, 3, 5 and 7 had high homology among themselves,
'outside ofthis 525 by region (Figures 13 (A) and (B)).. ~ . '
(00184] The following fragments of the members of the RENT family,
including the SEQ ID NO:l, have been characterized, and their utility
demonstrated
herein. For example, the fragment comprising nucleotides: '
1660-1992 (-394 to -62 fragment) enhances expression of the -46 minimal.
promoter of 355, and a fragment of T1275 (see Bstl-GUS; Bstl-355, Bst2-GUS,
Bst2-3JS, of Ir'lgulG JD);
1660-1875 (BstYI-DraI fragment; see Figure SC; -394 GUS-nos; and Table
6) The data in Table 6 indicates that this fragment acts as an enhancer;
1660-2224 (BstYI-SmaI fragment; see Figure SC; -394-GUS-nos) The
activity of this fragment is described in Tables 5 and 6;
1724-2224 (Figures 13C, and Figure 14A, tCUP RENT);
1875-2086 (DraI-NdeI fragment; core promoter element), see Figure SC
and Table 6 (-197-GUS-nosy;
1875-1992 (DraI -62 fragment) This fragment is shown to enhance
expression of the -46 minimal promoter of 355, and a fragment of T1275, as
shown in
82
CA 02507563 2005-05-13
Figures 5D'(see Dral-GUS; Dra2-GUS; Dral.-355; Drat-35S), and Figures SE-.G
(Drat-355; Drat-35S), and functions as a transcriptioral enhancer; . ' ,
2084-2224 (NdeI-SmaI fragment; or "N"; fiables 10-12, Figure S1B (+30-
. ,,
GUS-nosy, Figure 7 (T1275-GUS-nos; 355-GUS-nosy, and Figure 11 (3,55+N-GUS-
.
nosy; ~ , '
~ , , ,
,, ,
2091-2170 (L1N) see'Tables 10-12.' ~ , ~ '
,.
'. ~ , ~'
[00185] Based on sequence similarity using ~1CBI Blast 2 analysis (default ~ '
i ~. , .
parameters: blastn matrix, Lambda =1.37, Kf0.71 l, H=1.31), the fragmerits
identified
in above, exhibit from about 90~ to 98% identity to similar length fragments
of the
RENT sequences (SLQ ID ND's: 5-9)~~ ~ '
[00186] To verify the number of, repetitive elements in the region' giving
rise to
regulatory,element activity, more precise measurements were performed using
slot
blot hybridization. Slot blots were probed under conditions of high stringency
level a,s
used for the Southern blot (data not presented). 'These results indicate
that'a range bf °
approximately 10 to 43 copies of similar repetitive elements were esti~~~ated
per
haploid genome of N. tabacum. When the same slot blots were washed at lower
stringency, the same stringency as used during library screening, a range of
approximately 62 to 199 copies of similar repetitive elements were estimated
per
haploid genome. .
[00187] RNase protection assays and probes spanning both strands of the
combined tCUP and downstream sequence region, in the areas encompassing probes
5
to 8 (probe 5 was a 578 by BstYI-SmaI fragment; probe 6 was a 574 by RsaI-RsaI
fragment; probe 7 was a 244 by RsaI-RsaI fragment; and probe 8 was a 321 by
Rsal-
XbaI fragment) did not result in any protection in the repetitive region.
RNase
protection assays performed under these conditions has previously been shown
to
tolerate single mismatches by protection of non-identical sequences. This
suggests
that protected fragments may be detectable if members of the RENT family were
83
CA 02507563 2005-05-13
,
~ . ~ ~ , r
transcribef, at least for those elements that exhibit high sequence
similarity.
Examples of those elements which may be~detectable xre those hybridizing at
high ~ '
stringency on blots or those from which the downstream PCR clones originated.
A
~~ ,
lack of open reading frames was obseived within the RENT sequences. Together,
this
suggests a lack of coding capacity within the sequenced region.
[00188] Thus the tCUP cryptic, constitutive regulatory element is contained '
'.
within a moderately repeated repetitive element, which is the first known
member of a ~ ~, , ,,
new repetitive element family. ' ~ ' ~ ' ,
,' , . ,
,;.
Prntp~l~ct ieplatinnt elP~trppCrat~nn ar"1 C,ul~,u~e
~,. ,
[00189] ~ 1 Plasmids, prepared as described above were amplified and isoiateil
to
produce a sufficient amount of DNA necessary for transient expression ix~ pea
protoplasts, using the Qiagen Plasmid ~VIidiKit (Qiagen Inc.). ,
[00190] Pea (Pisum sativum L. var. Laxton, Progress) seedlings were grown in,
soil at 18°C (16 hr light, 8hr dark; 15-20 p.mol rh 2 s') provided by
Philips~(USA) F2b '
T12'cool white' flourescent tubes and young (ally expanded leaves were
harvested
from 2-3 weeks old plants. Leaves surface sterilized 5 minutes in 5%
commercial
bleach (Javex) (1% NaOCI). The abaxial surface of leaves were gently rubbed
with
carborundum powder, rinsed three times with sterile water, midribs removed and
remaining leaf blade was cut by sharp razor into ~ca 1 cubic cni pieces and
floated
rubbed surface facing first enzyme solution containing 0.1 % (w/v) pectolyase
Y-23
(Seishin Pharmaceutical, Japan), 0.5% potassium dextran sulphate (Calbiochem,
USA) and 0.5 M mannitol (pH 5.5) and vacuum infiltrated for 15 minutes. The
leaf
tissues were then incubated at 26 °C for another 15 minutes on a shaker
at 60
excursions/min. The solution was then decanted by filtration through a 100
mesh
nylon filter and the remaining tissue was incubated for 1-1.5 hr in a second
enzyme
solution containing 1.0% (w/v) Cellulase Onozuka R-10 (Yakult Honshu, Japan),
Pectolyase Y-23 0.05% (w/v) (Seishin Pharmaceutical, Japan and 0.5 M mannitol,
pH
6.0 at 26 C with 60 excursions/min.
84
CA 02507563 2005-05-13
[00191 ] The protoplasts were collected by filtration through a 100 ~.m nylon
mesh filter followed by centrifugation at 500 rpm for 5 min. The protoplasts
were,
gently collected by a wide bore pippet and washed twice with WS
electroporation
' buffer ( 4.Sg NaCI, O.Sg glucose, 9.2g CaCl2 , 2.Og KC in 500 ml) for 5 min
at 500
rpm and finally suspended at approximately 1x106/ml in 0.5 M m~nnitol
containing
150 ~,M MgCl2.
[00192] The viability of protoplasts was confirmed by FDA (Fluorescein
diactate) and alanine blue staining and protoplasts were kept on ice for 30
'minutes
prior to electroporation. A 25-30 u.g luciferin and desired DNA was added to
500;!! '
protoplast suspension, mixed gently and elecr_roporated at 100 uF and 200 v
Lsi_r._g
Gene Pulser II (BioRad). The electroporated protoplasts were kept on ice for 1
s-30
min, centrifuged for 5 min at 500 rpm and mixed with 0.5 ml growth medium. The
cultures were kept in dark at 25°C for 24 hr.
[uv i 93] Te each 500~i of protopiast suspension 200 ~.i of buffer solution
containing 100 mM KP04, 1mM EDTA, 10% glycerol, 0.5% triton x-100, 7 mM [3-
merceptoethanol was added and protoplasts were lysed and 1»ciferase and GT TS
activities were measured as described in Jefferson 1987 and Mathews et al.,
1995
(Jefferson, R.A. 1987. Assaying chimeric genes in plants: the GUS fusion
system.
Plant Mol. Biol. Reporter 5:387-405; Mathews, F.B., Saunders J.A., Gebhardt
J.S.,
Lin J-J., and Koehler M. 1995. Reporter genes and transient assays for plants.
In "
Methods in Molecular Biology, Vol 55: Plant Cell Electroporation and
Electrofusion
Protocols" ed. J.A. Nickoloff Humana Press Inc., Totowa, N.J. pp.147-162). All
GUS
activities were normalized with respect to luciferase activities to account
for variation
caused by electroporation.
[00194] When RENT sequences were cloned and tested for GUS transient gene
expression, all RENT sequences demonstrated high regulatory element activity
(Figure 14 (A)).
8~
CA 02507563 2005-05-13 '
.
s
[00195] ~ Figure 14 (A) shows that each.of the~regulatory elements isolated
from .
1
the 5 RENT sequences (RENT 1, 2, 3,.5, 7 'and tCUP-RENT) is capable of driving
the
expression of a coding region of interest (ix~ this case GUS) with which they
are in
~ , i~ n , I ' ' 1
operative association. The RENT regulatory elements resulted in more GUS
activity'
than that observed with .the 35S~minimal promoter-GUS construct (35S-46;
Figure 14
(A)). Furthermore, these results demonstrate that the RENT regulatory
sequences are ~ '
1
active in a heterologous species (pea).
'~'
~. ~ . , , r
Constitutive e~ ne expressibn by -394t-CUP seguenc~ in fraris~enic Arabidopsis
1 ,,
thaliana L. , ~ , '
[vvi 9v] , ~iiu''uiu'tip~is i~iaiiana (e,cotype Co~iumbiaj was grown in a
growth ,
chamber (16 hr of light and 8 hr of darkness at 23°C). Plants with
immature floral
buds and few siliques were dipped into, a solution containing Agrobacterium ~
~ ,
tumefaciens, 2.3 g/L MS salts (Sigma), 5% (w/v) sucrose and 0.03% Silwe~ L-77
(Lehle Seeds. Round Rock; TXl for 0-S _m__,'_n_ T1 CPPdc ~7VPrP rottPct~,d~
d,.ie~ zt 2c°C;
..
and sown on sterile media containing 40qg/mL kanamycin to select the
trahsformants.
Surviving Tl plantlets were transferred to soil and used for the analysis of
GUS
activity. For histochemical GUS assay, tissue was incubated in a 0.5 mg/ml
solution ,
of 5-bromo-4-chloro-indolyl 13-D-glucuronide in 100 mM sodium phosphate
buffer,
pH 7.0, infiltrated in a vacuum for half a hour and incubated at 37° C
overnight.
Following the incubation, tissue was washed in 7.0% ethanol to clear off
chlorophyll.
[00197] Arabidopsis plants were transformed with -394t-CUP-GUS fusion
gene. This fragment of tCUP exhibits substantial homology with the other
identified
RENT sequences (Figure 13 (B). The result, presented in Figure 14 (B),
demonstrates
that the -394t-CUP sequence drive constitutively GUS reporter gene expression
in all
organs such as leaves, stem, roots, and floral organs in transgenic
Arabidopsis. Since
this region is common to the characterized RENT sequences these results
indicate that
all RENT sequences contain regulatory elements capable of regulating
constitutive
gene expression.
86
CA 02507563 2005-05-13
Activity of the T1275 Regulatory Element ,
[00198) Analysis of leaves of randomly-selected, greenhouse-grown plants ~ ,
regenerated~from culture revealed a wide range of GUS specific activities
(Figure 6 '
(A); T plants). Plants transformed with pBI 121 (CLONETECH) which contains the
3~S-GUS-nos gene yielded comparable specific activity levels (Figure 6 (A); S
plants. Furthermore, the GUS protein levels detected by Western blotting were
similar between plants transformed with either gene When the GUS specific
activities
. were similar~(Figure. 6 (C)).
,,
' [00199) Generally, the IGVeI Vf V~S mRi~A in the leaves as determined by '
R~Tase protection (Fig~,re 6 (B)) c";-,.~~~t~.a ...:,,-.~ r~.~~
v m.auwu w1 1 llle Vvu SpeCIIIC aCtiVlIleS, however,
the level of GUS mR~~IA was about 60 fold (mean of 13 measurements) lower in
plants transformed with the T1275-GUS-nos gene (figure 6(B)) when compared
with
plants transformed with 35S-GUS-nos.
[uU2ii0) Since the levels of protein and the activity of extractable protein
were
similar in plants transformed with T1275-GUS-nos or 35S-GUS-nos, yet the mRNA
levels were dramat,'_cal_ly different, these results suggested the existence
of a regulatory
element downstream of the transcriptional start site in the sequence of T1275-
derived ,
transcript.
Post-Transcriptional Regulatory Elements within T1275
[00201 ) An experiment was performed to determine the presence of a post-
transcriptional regulatory element within the T1275 leader sequence. A portion
of the
sequence downstream from the transcriptional initiation site was deleted in
order to
examine whether this region may have an effect on translational efficiency
(determined by GUS extractable activity), mRNA stability or transcription.
[00202) Deletion of the Ndel-Smalfragment ("N"; SEQ ID N0:2) from the
T1275-GUS-nos gene (Figure 15; T1275-N-GUS-nos; includes nucleotides 2084-
2224 of SEQ ID NO:1) resulted in at least about 46-fold reduction in the
amount of
87
CA 02507563 2005-05-13
~~ , i
GUS specific activity that could be detected in leaves of transgenic tobacco
ev ,
,
Delgold (see Table 7). Similar results, of about at least a 40 fold reduction
in GUS .
. ., ,
activity due to the deletion of the Ndel-Smal fragment, were observed in
transgenic
~, ,
tobacco cv SRl and transgenic alfalfa (Table 7)'. , Addition of the same
fragment
(Ndel-Smal) to a 35S-GUS-nos gene (Figure 7; 35S+~I'-GUS-nos) construct
increased ,
the amount of GUS specific activity by about 5-fold. in tobacco, and by a much
higher '
amount in alfalfa (see Table 7). ~ '
' '
Table 7: GUS specific activity in leaves of greenhouse=grown transgenic
tobacco ' .
cv Delgold, SRl and transgenic alfalfa tra~isformed with vectors c~esigned~to~
assess the presence of cryptic regulatory sequQnces within the transcribed
sequence derived from the T12'S~ G>.1S gene fusion (see Figure 7): MeantSE(n).
Construct , GUS specific activity ~ ~ ,
pmoles MU/min/mg protein ;
Delgold (1)'Delgold (2) ~ SR1 Alfalfa
T i275-GUS-nos557ti83 (2i)493ti57 (25)~ 805+253 (22) 18~7t64
(24)
T1275-N-GUS- 1213 (22) 1213 (27) ' , 612 (25) '40.5 (25)
~ ~
nos
35S-GUS-nos 1848=692 i347~ 41 S (26) 1383+263 (25) 1711
(15) (24)
35S+N-GUS- 69903148 6624.12791 (26) 619211923 1428601
(23) (24)
nos (24)
[00203] A similar effect was noted in organs tested from transformed tobacco
(Table 8) and alfalfa plants (Table 9)
88
CA 02507563 2005-05-13 '
Table 8: Expression of T1275-GUS-nos (+N) compared with T1275-(-N)-GUS-
nos (-N) in organs of transgenic tobacco cv. Delgol~and SRl. Mean~SE(n=5).,
Organ GUS specific Activity (pmol MU/min/mg/protein)
Delgold ~ , SR1
+N -N +N -N
Leaf, '1513+222 35+4~ 904138 4~1 ,
Flower 360+47 38+8 ~ 175+44 28+3 '
Seed 402+65 697 ' 370+87~. 335
' , ,
Table 9: Expression of T1275-GUS-nos, T1275-I-N)-GUS-nos; 35S-~I tS-n_r~z;
35S-GUS(+N)-GUS-nos in organs of transgenic alfalfa. Mean~SE(n=S).
Construct GUS Specific Activity (pmol Mu/min/mg protein) .
Leaf Petiole Stem Flower
~~.u- v.~ i.~;~~~.~.~ trL,tW-iG.l 1.J00./~GbU 4jb.1~16U.y
T1275(-N)GUS 5.411.4 7.6+1.2 8.12.0 7.2511.7
35S-GUS 67.550.3 48.9+23.2 56.8128.7 23.217.3
35S(+N)GUS 5545+2015 107916194 9931+5496 1039+476.7
Control 3.7 13.2 11.8 18.7
[00204] In transient expression assays using particle bombardment of tobacco
leaves, the Nde 1-Sma 1 fragment fused to the minimal -46 3 5 S promoter
enhanced
basal level of 35S promoter activity by about 80 fold (28.67 X2.91 v. 0.33
+0.33
relative units; No.blue units/leaf).
[00205] SEQ ID N0:2 comprises nucleotides 2084 to 2224 of SEQ ID NO:1.
Nucleotides 1-141 of SEQ ID N0:2 comprise nucleotides obtained from the plant
portion of T1275 (nucleotides 2084 to 2224 of SEQ ID NO:l). Nucleotides 142-
183
of SEQ ID N0:2 comprise vector sequence between the enhancer fragment and the
GUS ATG. The GUS ATG is located at nucleotides 186-188 of SEQ ID N0:2.
89
CA 02507563 2005-05-13
[00206] ~ A shortened fragment of the Ndel-SrreaI fragment (see SEQ ID N0:3),
referred to as "ON", "dN", "deltaN" or "tCUP delta" and lacking the out~of
frame ,
,, ,
upstream A'TG at nucleotide 2087-2089 of SEQ ID NO:l, was also constructed and
tested in a variety of species. ON was~created by replacing the NdeI site
(Figure 10(A)) ,
within the leader sequence to a BgllI site thereby eliminating the upstream
ATG at
position 2087 of SEQ ID NO:1. A Kozak consensus sequence was also constructed
at
the initiator MET codon and a NcoI site was added to facilitate construction
with other , ,
coding regions (see Figure 10 (B)). Nucleotides 1-86 of SEQ ID N0:3 (i.e. ~N
with
Ko'zack sequence) are derived from T1275 (nucle'otides 2086-2170 of SEQ ID
NO:1).
L1N also includes a Kozack sequence ,from nucl_eotid'es 87 to 97 of SEQ ID
?10:3, ar~d
nucleotides 98 to 126 of SEQ _ID N0:3 comprise thewector Seq»e_n_ce her~.uee.n
the
enhancer fragment and the GUS ATG. The GUS ATG is located at nucleotides 127-
129 of SEQ ID NO:3. '
' [00207] Constructs comprising AN, for example T1275(~N)-GUS-nos, ,when ,
ii~urv~i.'W~eV iritV tVVClvIiV ylCldGd J foil greater ieveis of GUS activity
in leaves of
transgenic tobacco (5291986 pmolMU/minlmg protein; (n=29) compared to plants
expressing T1275-GUS-nos (1115299 pmol MU/min/mg protein, n=291.
Activity of NdeI-Smal, N, and ON in other species '
[00208] In monocots, transient expression in corn callus indicated that the
NdeI-SmaI fragment (SEQ ID N0:2), or a shortened NdeI-SmaI fragment, ON (SEQ
ID N0:3), significantly increases GUS expression driven by the 35 S promoter,
but
not to the higher level of expression generated in the presence of the ADH1
intron
("i"; Figure 11 and Table 10).
CA 02507563 2005-05-13
Table 10: Transient expression analysis of GUS activity in bombarded corn
calli. Luciferase activity was used to normalize the'data. Mean tse (n=5).
Construct Ratio GUS:Luciferase activity ~ ,
35S GUS-nos 7.4+4 ,
35S(+N)-GUS-nos 19+5 ~ , '
35S(ON)-GUS-nos 18+10 ~ ~ , ~ ' '.
35S-i-GUS-nos 66+27
,,
.. , ,
[00209] The ~nctionality of the t1%dei-SmaI fragment (SEQ ID N0:2) was also
deteriiiiiivu iii nvir-piallt JpV.V.iGJ. 111 l.Vill Grs, for~exdrrrpie white
spruce, transient
. ~. ~ ,
bombardment of cell culture exhibited an increase in expression (Table 11).
Table 11: Expression of T1275-GUS'-nos, T1275(-N)-GUS-nos, 35S-GUS-nos,
35S (+N)-GUS-nos in white spruce embryonal masses following bombardment
(n'3). , . ~ .
Construct Average GUS expression per leaf
(Number of blue s ots)
T1275-GUS-nos 72.6719.33
T1275(-N)-GUS-nos 21.33+4.49
35S-GUS-nos 113.67+17.32
35S(+N)-GUS-nos 126.33+19.41
* average spot much greater in size and strength.
[00210] In yeast, the presence of the NdeI-SmaI fragment (SEQ ID N0:2) or
~N (SEQ DI N0:3) exhibited strong increase in expression of the marker gene. A
series of constructs comprising a galactose inducible promoter Pg~~, various
forms of
the Ndel-Smal fragment, and GUS (UidA) were made within the yeast plasmid
pYES2. A full length Ndel-Smal fragment N (pYENGUS), dN (containing a Kozak
consensus sequence; pYEdNGUS), and ~N without a Kozak consensus sequence
(pYEdNMGUS; or ONM) were prepared (see Figure 12, and SEQ ID N0:4).
91
CA 02507563 2005-05-13 .
[00211] ~ Nucleotides 1-86 of SEQ ID N0:4 (~NM ) comprise a portion of the
enhancer regulatory region obtained from T1275 (n~icleotide 2086 -2170'of SEQ
ID
,,
NO:1 ), while nucleotides 87-116 comprise a vector sequence between the
enhancer
' ' ' fragment and the GUS ATG which is.located at nucleotides 117-119 of SEQ
ID N0:4. , ~
,,
~ ' ~ [00212] These constructs were tested in yeast.strain INVSCI using known
transformation protocols (Agatep R. et al. 1998; bioriiednet.com/db/tto): The
yeast
were grown in non-inducible medium comprising raffinose as a carbon source for
48hr at 30°C' and then transferred onto inducible medium (galactose as
a carbon
,
source). Yeast cells were harvested after 4 hr post induction and GUS activity
determined quantitatively. Up to about a 12 fold inc~'ease in activity was
observed
with constructs comprising ON. Constructs comprising ONM exhibited even higher
levels of reporter activity. The results indicate that the Ndel-Smal fragment
(SEQ 11'7 '
N0:.2), ON (SEQ ID N0:3) and ONM (SEQ ID N0:4) are functional in yeast (Table
~
12). ~ , ,
Table 12: Expression of pYEGUS, pYENGUS, pYEdNGUS, and pYEdNMGUS
(0N, without a Kozak consensus sequence) in transformed yeast (n=5).
Construct Expt. 1 Expt. 2 ,
Activity Activity
pYES-GUS-nos 9315 407+8
pYES(+N)-GUS-nos 753186 1771191
pYES(ON)-GUS-nos 1119+85 2129+166
pYES(ONM)-GUS-nos 173145 6897536
[00213] Constructs containing ONM (i.e. ON lacking the Kozack sequence; SEQ
ID N0:4) were also tested in insect cells. These constructs comprised the
insect virus
promoter ie2 (Theilmann D.A and Stewart S., 1992, Virology 187: pp. 84-96) in
the
present or absence of ONM and CAT (chloramphenicol acetyl-transferase) as the
reporter gene. The insect line, Ld652Y, derived from gypsy moth (Lymantria
dispar)
92
CA 02507563 2005-05-13 '
,~ , , ;
was transiently transformed with the above' constructs using liPosomes
(Campbell
M.J. 1995,.Biotechniques 18: pp. 1027-1032; Forsythe LJ. et al 1998, Virology
252:
. , ,
pp. 65-81 ). .Cells were harvested 48 hours after transformation and CAT
activity
,a
quanitatively measured using tritiated acetyl-CoA (Leaky P. et al: 1995
Biotechnique~
19: pp. 894-898). The presence of the transl,ational enlz'ancer was found to ~
~ I~ ~ , '
significantly modulate the activity of the insect promoter-reporter gene
construct in '
. , , ,
insect cells. ~ ' ~ ' '
~ I
y
[00214] ' Bacteria we're transformed with either'pBI22'1, comprising 35S ,
promoter and GUS, or 35S-N-GUS , comprising the full~length Ndel-Smal fragment
(SEQ ID N0:3). Since uidA (GUS) is native to ~.coli, two uidA mutants, uidAl
and
, , '
uidA?, that do not express uidA, w~re'used for these experiments (mutants
obtained
from E.coli Genetic Center 335 Osborn Memorial Laboratories, Department of
Biology, Box 208104, Yale University; New Haven CT 06520-8104). These bacteria
' ,
were transformed using standard protocols, and' tr~nsformants were assessed by
assaying GUS activity from a 50 ~.l aliquot of an overnight culture. The "N"
fragment
,~ .
(35s-N-GUS) was observed to modulate the activity of the reporter gene in
bacterial
cells.
[00215] These data are consistent with the presence of a post-transcriptional
regulatory sequence in the NdeI-SmaI fragment.
The Ndel Smal fragment functions as a transcriptional enhancer or mRNA
stability determinant
[00216] The levels of mRNA were determined in leaves obtained from plants
transformed with either T1275-GUS-nos, T1275-N-GUS-nos, 35S-GUS-nos, or
35S+N-GUS-nos (Figure 9 (A)). Relative RNA levels were determined by
ribonuclease protection assay (Ambion RPAII Kit) in the presence of a-32P-CTP
labeled in vitro transcribed probe and autoradiographic quantification using
Kodak
Digital Science 1D Image Analysis Software. Hybridization conditions used
during
93
CA 02507563 2005-05-13
RNase protection assay were overnight at 42-45 degrees in 80% formamide, 100
mM
sodium citrate pH 6.4, 300 mM sodium acetate pH 8.~, 1 mM EDTQ. ~ , ,
,,
X00217] The levels of mRNA examined frorr~ transgenic tobacco plants '
transformed. with either T127~-GUS-nos, T1275-N-GUS-nos, 35S-GUS-nos, or
35S+N-GUS-nos, were higher in transgenic plants comprising the~Ndel Smal
fragmgnt under the control of the T1275 regulatory element but lpwer in those
under
the control of the 35S promoter, than in plants comprising constructs that
lack this
region (Figure 7 (A)). This indicates that this region functions by either
modulating
'transcriptionai rates, or the stability of the transcripts, or both. ~ , '
~~le ~~lQ~ .~'mnT ~ra~_m__oj,t fn~n~f« ~ 2;; a ti au~i~a~~Tiai cyiiai3C~r'
[00218] Analysis were perforrzed in order to determine whether the Ndel Smal '
region functions post-transcriptionally. The GUS specific activity:relative
RNA level
was determined from the GUS specific activity measurements, and relative RNA .
ieveis in greenhouse grown transgenic plants (figure 9 (B)). The ratio of GUS
specific
activity to relative RNA level in individual transgenic tobacco plants
comprising the
Ndel-Smal fragment is higher than in plants that do not comprise this region
(Figure 9
(B)). Similar results are obtained when the data are averaged, indicating an
eight fold
reduction in GUS activity per RNA. Similarly, an increase, by an average of
six fold,
in GUS specific activity is observed when the Ndel Smal region is added within
the
35S untranslated region (Figure 9 (B)). The GUS specific activity:relative RNA
levels are similar in constructs containing the Ndel Smal fragment (T1275-GUS-
nos
and 35S+N-GUS-nosy. These results indicate that the Ndel-Smal fragment (SEQ ID
N0:2) modulates gene expression post-transcriptionally.
[00219] Further experiments, involving in vitro translation, suggest that this
region is a novel translational enhancer. For these experiments, fragments,
from
approximately 3' of the transcriptional start site to the end of the
terminator, were
excised from the constructs depicted in Figure 7 using appropriate restriction
endonucleases and ligated to pGEM4Z at an approximately similar distance from
the
94
CA 02507563 2005-05-13
I
' ~~ ' , '
transcriptional start site used by the prokaryotic T7 RNA polymerase. Another
' I
construct containing the AMV enhancer in the 5' UTR' of a GUS-nos fusion was ,
similarly prepared. This AMV-GUS-nos construct was created by restriction
,
endonuclease digestion of an AMV-GUS-nos fusion, with Bglll and EcoRl, from '
pBI525 (Dada et al., 1993, Plant Science 94; 139-149)~~and ligation with
pGEM4Z , '
(Promega) digested with BamHl and EcoRl. Transcripts were prepared in vitro in
the ~ '
resence of m~G 5' S' G Ca Analo Ambion . Transcri is were translated in
P ( )PPP( ) P g ( , , ~ p '
vitro in Wheat Germ Extract (Promega) in the presence,of 35S-Mexhion~ne and
fold ~ ~ '~
I ~ ,
enharace_m__ent calcol_atPd from TC_'_A_~prPE,i_Yi_tahl'P rp~n~c , ,
[00220] Translation of transcripts in vitro, demanstrate an increase in
transiational efficiency of RhlA containing the Ndel to Srizal fragment (see
Table'13).
Table 13: In vitro translation of mRNA obtained from transgenic tobacco plants
,
transformed with vectors with or without a Ndel-Smal fragment obtained from
the T127S f 1TC gPnP fn_cinn (CPP FigyrP 7), ycins ;xy~~t grr ~.. ~..~.-/] ~
1t 1111 ~\.H I
.
' I iI
in vitro translation
in vitro transcriptfold Qnhuncement
T1275-GUS-nos 3.7
TI275-N GUS-nos1
AMV GUS-nos 1.9
[00221 ] The levels of protein produced using mRNAs comprising the NdeI-
SmaI fragment are also greater than those produced using the known
translational
enhancer of Alfalfa Mosaic Virus RNA4 (Jobling S.A. and Gehrke L. 1987,
Nature,
vol 325 pp. 622-625; Datla R.S.S. et al 1993 Plant Sci. vol 94, pp. 139-149).
These
results indicate that this region functions post-transcriptionally, as a
translational
enhancer.
CA 02507563 2005-05-13
, ,
[00222] All citations are hereby incorporated by reference. The nucleic acid
, ,
sequences fisted in the Sequence Listing filed herewith are incorporated by
reference
into this application in their entireties. ~ ~ , ,
,, , ,
,,
[00223] The present invention has been described with regard to ,one or mole.
~~
embodiments.. However, it will be apparent to persons skilled in the art that
a number ,
of variations and modifications can be made without departing from ~he~ scope
of the ' ',
invention as defined in the claims.
I ,, , , ,
WH_A_T TC f_'T,_A_T__.M_F.TI TC~ ~ '
- ~ , , , , ,
96